Почему лил чсв: «День, когда меня кикнули, был одним из самых худших в моей жизни, сегодня же — один из лучших»
«День, когда меня кикнули, был одним из самых худших в моей жизни, сегодня же — один из лучших»
Участник состава Winstrike Team Илья «Lil» Ильюк прокомментировал поражение своих экс-тиммейтов на The International 2019. Ильюк вспомнил о том, как его выгнали из Virtus.pro, и обвинил свою бывшую команду и организацию в клевете, попытках испортить ему жизнь и сломать карьеру.
Сразу предупреждаю тех, кто пришел за зрелищем – здесь его не будет. Ровно так же как и не будет оскорблений, обидок, перехода на личности и любого негатива по отношению к кому-либо. Просто мысли и сухие факты.
Как вы знаете, сегодня Virtus.pro вылетели с инта, заняв при этом 9-12 место.
Я рад. И рад я не тому, что кто-то выиграл, а кто-то проиграл. А тому, что это все наконец закончилось.Полтора года назад меня кикнули из вп. Не буду скрывать, что я продолжительное время таил обиду на ребят, также я согласен с тем, что где-то сам был неправ, где-то перегнул, да и вообще лучше бы наши пути просто разошлись мирно.
Жаль этого не случилось. А случилось то, что аффектит меня до сих пор. Последние полтора года были самыми сложными в моей карьере, во многом из-за всего того, что на меня вылилось после кика со стороны моей бывшей команды и организации. Со мной никто не хотел ни играть, ни иметь что-либо общее. И вовсе не потому что я плохой_игрок или не_такой_человек. А только потому что у всех появились ложные предубеждения по отношению ко мне из-за высказываний некоторых личностей. Очевидно, что когда ты на коне, тебя все послушают и примут твои слова за истину. Делалось это чтобы любым способом отбелить свою репутацию, ведь какое кому дело до какого-то там челика, подумаешь карьеру ему сломаешь (почти). Мне не хватит символов, чтобы процитировать все высказывания и всю ложь, всю клевету, которая была сказана в моей адрес (и про медийку, и про личную жизнь, и про отношение к тренировкам, про сложный характер, про плохой уровень игры, про психологические проблемы и т.д и т.п.), или таким образом, чтобы испортить мне жизнь. И не только в соц.сетях, а и в личных разговорах. Не хочу больше никогда вспоминать и возвращаться к этому. Жизнь продолжается, а жить с этим грузом внутри я не хочу, и давно не живу. День, когда меня кикнули, был одним из самых худших в моей жизни, сегодня же один из лучших. Сегодня всем должно стать понятно, что было правдой, а что нет. В любом случае, спасибо ребятам за все эмоции, они большие молодцы и проделали огромную работу, за это респект. И самое главное – спасибо за безмерную мотивацию не останавливаться все это время несмотря ни на что. Мало что в этой жизни заставило бы меня просыпаться в 4 утра, чтобы посмотреть дотку.
Полтора года назад, когда узнал новость о своем расставании с командой сидя на буткемпе, последнее, что я услышал было «Мы не выиграем с тобой инт». Ну что же, без меня вы тоже его не выиграете 🙂
Это мой гвоздь в крышку гроба всей этой ситуации.

Генеральный менеджер Virtus.pro Роман Дворянкин ответил на заявление Ильюка.
Во втором раунде лузеров Virtus.pro всухую проиграла коллективу Royal Never Give Up под предводительством Zhang «LaNm» Zhicheng. Команда получила в награду $675+ тысяч призовых.
Генеральный менеджер Virtus.pro Роман Дворянкин подвел итоги сезона и прокомментировал неудачное выступление команды на главном чемпионате года в Шанхае. Комментарий Владимира «No[o]ne» Миненко вы найдете в нашей новости.
Смотри The International 2019 с Dota2.net
Источник: t.me
«Я никогда не был ЧСВ», сообщает Lil — Dota 2
Игрок Natus Vincere Илья «Lil» Ильюк рассказал о своей жизненной позиции.
«Я всегда был реалистом, им же и остаюсь. Вижу мир таким, какой он есть, не лучше – не хуже, без всяких призм. Благодаря тому, что я очень редко руководствуюсь эмоциями – я могу трезво смотреть на вещи и ситуации, адекватно оценивать свои силы, возможности. Ко всему прочему я очень прямолинейный человек, всегда говорю, что думаю/чувствую в конкретный момент времени.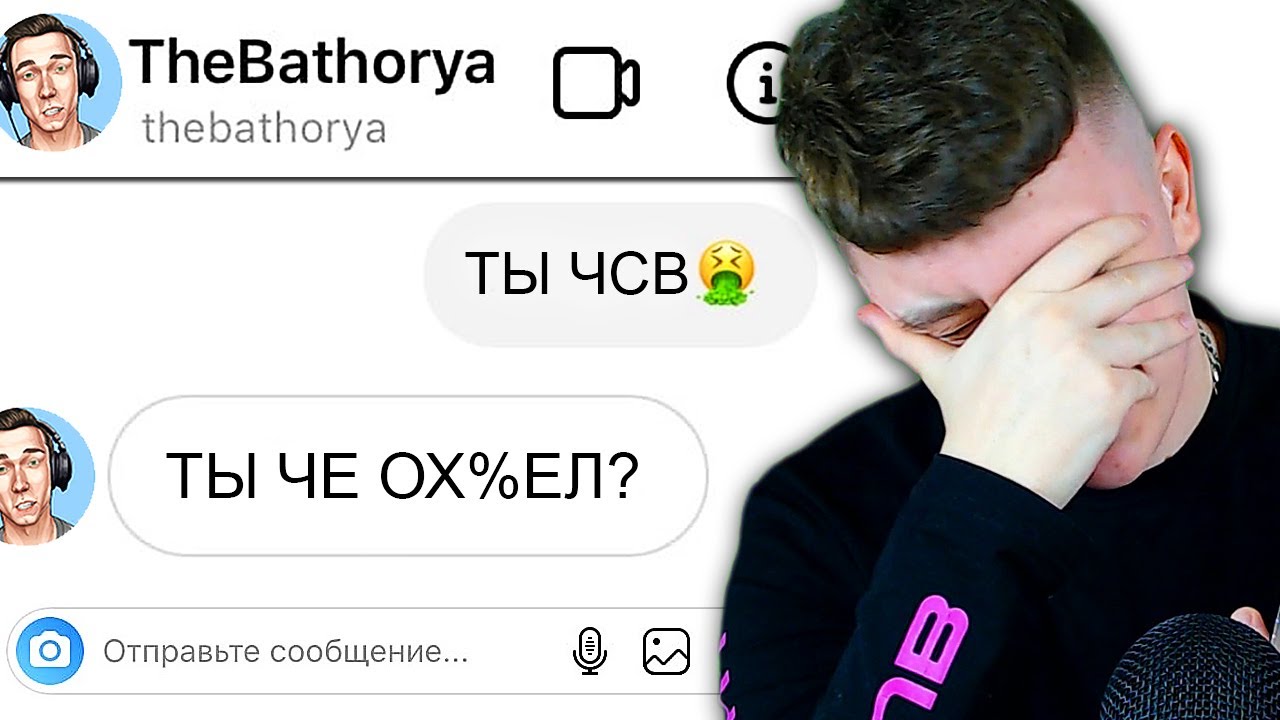 Я не жалуюсь и не ищу поддержки, не прошу понимать меня.
Я не жалуюсь и не ищу поддержки, не прошу понимать меня.
К чему я это все. В последний месяц моя жизнь существенно изменилась и заиграла новыми красками (преимущественно черно-желтыми), и те, кто следят за моей «деятельностью» всецело или, по крайней мере, за инстаграмом, наверняка заметили изменения в том, что я говорю, и как я это преподношу. Куда же делся тот Чрезвычайно Самоуверенный Выскочка, у которого все «легко» и который заставляет «сушить весла» или бьет всем «по губам»? Дак вот его и не было никогда, и образа не было. Просто совокупность мыслей и чувств в конкретный момент времени в несколько шуточной форме.
Возвращаясь к теме команд и моих взаимоотношений с людьми… Из-за того, что я сам по себе достаточно мягкий и добрый человек по отношению к людям, которых я считаю «близкими», я всегда пытался понять и подстроиться для всеобщего блага, по крайней мере, мне так казалось. В конечном счете люди воспринимали мою «доброту» за слабость, «садились на шею», «взвешивали ножки», пытались подавлять, ну и дальше мы знаем чем обычно это все заканчивается.
Как показывает практика, такой тип поведения, которого я придерживался, на дистанции абсолютно не работает, и, наверное, в какой-то степени здорово, что я сейчас это определил, хоть и таким «болезненным» образом. А так как выводы сделаны, сейчас я могу все изменить в самом начале, сделать все по-другому и я приложу больше максимума усилий в рамках своих возможностей для этого. Для себя и для команды.
Это невероятно сложно – начинать все с нуля, но я буду пытаться изо всех сил», – написал Лил в инстаграме.
Лил забросил Пая на горку
У Na`Vi сразу три крупных проблемы
AbitLazy – Наши Rhymes (feat. ЧСВ Минус) текст и слова песни
Слова
[Припев]:Мама мне сказала то, что рэп — кал, рэп — кал
Батя микрофон мой спиздил и сказал, что я еблан
Мы достойны премии начала, дольше всех начал
Полтора года нас слушал только FAT MOON и я
А я буду делать рэп, ведь ниче не умею больше, будем вместе делать деньги и трясти цепями, точно, знаю, мы подольше потянем, останусь вечно молодым и вечно пьяным
Мне кричит мой препод, что я хули-хулиган
Моё желание — сунуть его в улей и капкан
Вы хотели свэга или может быть похаслить
Мы пойдем обедать, ведь мы очень устали
Ты понял?
Не ври, ты нихуя не понял, сука
Вот в этом и прикол, прикинь? Тополиный пух в июле
Похуй, что ты слушаешь.
 Главное — качает
Главное — качаетСлышишь что мы в уши льем?
Чувствуешь, воняет?
[Чсв минус part]:
Чсв плюс? Нет, чсв минус!
Я подвинусь? Ха, нож в спину!
Все твои заслуги — всего лишь формальность
[FAT MOON part]:
Я съел деда, мой пёс сгрыз бабку
Они говорили то, что рэп — кал
Я отобедал, пёс порвал как тряпку
За рэп и двор кидаем в мангал
[Припев]:
Мама мне сказала то, что рэп — кал, рэп — кал
Батя микрофон мой спиздил и сказал, что я еблан
Мы достойны премии начала, дольше всех начал
Полтора года нас слушал только FAT MOON и я
А я буду делать рэп, ведь ниче не умею больше, будем вместе делать деньги и трясти цепями, точно, знаю, мы подольше потянем, останусь вечно молодым и вечно пьяным
[Bridge]:
Мы не курим, мы не курим, но под кайфом мы внатуре х4
[FAT MOON part]:
Это хип-хоп, а рок идёт в жеппу
Русский реп говна хлебнул и взобрался к небу
То, что так всё поменялось — это круто
Прошли те времена, когда писали рэп — кал
Этот ссаный рок актуальность потерял
Правда в репе много лалок, такого положение
Но мы хотя бы делаем годное сведение
[Чсв минус part]:
Лучше делать рэп, чем раздавать листовки —
Об этом ты подумаешь, стоя на остановке
Увидев в чёткой тачке меня у штурвала
Поймёшь не подарили, а начитала
[Припев]:
Мама мне сказала то, что рэп — кал, рэп — кал
Батя микрофон мой спиздил и сказал, что я еблан
Мы достойны премии начала, дольше всех начал
Полтора года нас слушал только FAT MOON и я
А я буду делать рэп, ведь ниче не умею больше, будем вместе делать деньги и трясти цепями, точно, знаю, мы подольше потянем, останусь вечно молодым и вечно пьяным
[Lil d part]:
Зачем писал эти биты, зачем назвал себя Lil d?
Просто я вижу успех, потенциальны наши рты
Кто-то говорит, что рэп тупее воды, но
Пропоганду мы распространим через фиты
[FAT MOON part]:
Рок сасать, мы на коне
Репчик можно записать на любом говне
И не нужен самый жоский микрофон
Чтобы зачитать что хейтер — гандон
Аасамаламалейкум моим братьям, детям, тем, кто в репе
Думаешь что жоский ты, но на бите мы, блять, свирепы
Нужно обосрать всех в треке, чтобы были целы скрепы
Вы сказали шарите за рэп, но вы просто слепы
[Чсв минус part]:
Мама говорит мне: иди учи уроки
Миру не нужны твои рэперские строки
Но crazy abitlazy щас со мной на блоке
И к чёрту это всё, мне плевать на их упрёки
Чекай наши rhymes, bitch, чекай наши rhymes, bitch х2
Моргенштерн по ящику? (Powerpuff не в Кащенко)
Вы возьмёте за щеку
Мой флоу с ужасной жаждою —
Его понять не каждому
Ведь я такая важная
ЧСВ – откуда пошло и на сколько люди ЧСВшные? | ЛюбопытNO
ЧСВ – это сокращение от выражения «чувство собственной важности» — эти слова встречаются в книгах Карлоса Кастанеды – известного мистика 20го века, познакомившего мир с тайными ритуалами и обрядами индейских шаманов.
ЧСВ, проще говоря, – это чувство превосходства над кем-то или чем-то, это оценка себя и своих поступков как чего-то более важного, чем у других.
Мой знакомый увлекался философией и психологией, он часто рассказывал, как некие мистические действия или утверждения в книгах эзотерического направления являются просто способом подготовить свой разум для того или иного действия. Иными словами, шаманы, стуча в бубен, настраивают свой разум на определенный лад. О том же говорит и психология: у всего есть причина, восприятие мира человеком достигается тем или иным способом формирования сознания.
Он поведал мне о том, что чувство собственной важности является уникальным качеством человека, которое обязано быть, именно это чувство позволяет развиваться, однако, если его чрезмерно много, то происходит перекос в восприятии человеком окружения, и он начинает обижаться на правду, правдивые моменты считать ложью, он становится агрессивным и более беспокойным. Из всего этого вытекает множество других проблем, усугубляя положение такого человека.
Современное общество очень сильно завязло в излишнем ЧСВ, или просто горделивости, всего должно быть в меру, когда чего-то становится слишком много или мало в человеке, он начинает болеть, а психологические болезни проявляются в ненормально острых реакциях на действия и происходящее вокруг.
А вот когда мы спокойно реагируем на всё, когда нас не выводят из себя мелочи или более крупные проблемы, когда мы можем согласиться с глупостью окружающих, т.к. они такие же люди, которые ошибаются и ищут возможности решения своих проблем, тогда мы становимся самодостаточными и готовы жить полноценно.
————————-
Только человек способный признаться себе в том, что он дурак, может развиваться и идти вперёд. Когда человек сможет признаться в своей глупости окружающим, не ожидая от них сочувствия или помощи, тогда он сможет жить в гармонии с самим собой и со всеми вокруг.
Если вам понравилась статья нажмите палец вверх, я увижу вашу реакцию и буду вам благодарен. Оставляйте свои комментарии и делитесь статьёй с вашими друзьями. Подписывайтесь на канал.
Оставляйте свои комментарии и делитесь статьёй с вашими друзьями. Подписывайтесь на канал.
Virtus.pro вылетела с The International 2019 — Киберспорт на DTF
Royal Never Give Up всухую обыграла Virtus.pro во втором раунде нижней сетки The International 2019.
RNG сыграет с победителем матча Liquid – TNC.
Virtus.pro заняла 9-12-е место и заработала более 674 тысяч долларов.
The International 2019 проходит в Шанхае с 15 по 25 августа. 18 команд разыгрывают более 33 млн долларов.
18 команд разыгрывают более 33 млн долларов.
5914 просмотров
{ «author_name»: «GERBARIY», «author_type»: «self», «tags»: [], «comments»: 161, «likes»: 79, «favorites»: 1, «is_advertisement»: false, «subsite_label»: «esport», «id»: 65139, «is_wide»: true, «is_ugc»: true, «date»: «Wed, 21 Aug 2019 14:27:43 +0300», «is_special»: false }
{«id»:3274,»url»:»https:\/\/dtf. ru\/u\/3274-gerbariy»,»name»:»GERBARIY»,»avatar»:»a89abf8f-6191-c106-9bf6-d496c757c423″,»karma»:2567,»description»:»»,»isMe»:false,»isPlus»:false,»isVerified»:false,»isSubscribed»:false,»isNotificationsEnabled»:false,»isShowMessengerButton»:false}
ru\/u\/3274-gerbariy»,»name»:»GERBARIY»,»avatar»:»a89abf8f-6191-c106-9bf6-d496c757c423″,»karma»:2567,»description»:»»,»isMe»:false,»isPlus»:false,»isVerified»:false,»isSubscribed»:false,»isNotificationsEnabled»:false,»isShowMessengerButton»:false}
{«url»:»https:\/\/booster.osnova.io\/a\/relevant?site=dtf»,»place»:»entry»,»site»:»dtf»,»settings»:{«modes»:{«externalLink»:{«buttonLabels»:[«\u0423\u0437\u043d\u0430\u0442\u044c»,»\u0427\u0438\u0442\u0430\u0442\u044c»,»\u041d\u0430\u0447\u0430\u0442\u044c»,»\u0417\u0430\u043a\u0430\u0437\u0430\u0442\u044c»,»\u041a\u0443\u043f\u0438\u0442\u044c»,»\u041f\u043e\u043b\u0443\u0447\u0438\u0442\u044c»,»\u0421\u043a\u0430\u0447\u0430\u0442\u044c»,»\u041f\u0435\u0440\u0435\u0439\u0442\u0438″]}},»deviceList»:{«desktop»:»\u0414\u0435\u0441\u043a\u0442\u043e\u043f»,»smartphone»:»\u0421\u043c\u0430\u0440\u0442\u0444\u043e\u043d\u044b»,»tablet»:»\u041f\u043b\u0430\u043d\u0448\u0435\u0442\u044b»}},»isModerator»:false}
Еженедельная рассылка
Одно письмо с лучшим за неделю
Проверьте почту
Отправили письмо для подтверждения
ЧСВ — что это значит, расшифровка в молодежной среде, стадии у человека, как ответить на сленг в ВК, играх.
 Примеры применения
Примеры примененияЛюбая информация воспринимается правильно лишь до того момента, пока известно значение любого из сказанных слов. При общении в любой среде, в том числе и молодежной, может понадобиться расшифровка значения неизвестного слова или сочетания букв. Таким неизвестным может стать аббревиатура ЧСВ.
Изречение известное большинству христиан – «Бог гордым противится!», имеет более глубокое значение. В греческом слово «гордый» звучит, как «гиперфания». В переводе с эллинского приставка «гипер» – «сверх», а корень «фания» – «явление».
Расшифровка слова гиперфания дословно – это показушность. Таким образом, Бог противится всем, кто выставляет себя, как лучшего в чем-либо. Такое поведение характерно для представителей молодежной среды, у которых большое ЧСВ.
Что такое ЧСВ
ЧСВ (расшифровка в молодежной среде звучит – Чувство Собственной Важности) является одним из модных сочетаний букв. Интернет-общение имеет свои особенности. Здесь нет возможности дополнить речь интонацией и мимикой. Зато есть некоторые всеми принятые краткие обозначения емких понятий. Контекст использования конкретной аббревиатуры, как правило, совсем недружественный.
Зато есть некоторые всеми принятые краткие обозначения емких понятий. Контекст использования конкретной аббревиатуры, как правило, совсем недружественный.
Сочетание буковок фигурирует в сюжетах тематических картинок, высмеивающих определенную категорию молодежи. Их можно встретить в язвительных комментариях. Используется аббревиатура в основном в молодежной среде.
Значение выражения часто путают с эгоизмом, но это разные характеристики.
Под воздействием ЧСВ теряется способность трезво оценивать ситуацию. Все происходящее воспринимается, с точки зрения болезненного эго. Большего распространения ЧСВ достигло в русскоязычном интернете. Его появление справедливо связывают с русским вариантом викиэнциклопедии Луркморье. Именно там впервые появился и стал популярным новый современный мем.
ЧСВ (расшифровка в молодежной среде иногда, может звучать, как Чувство Собственного Величия) имеет иностранные корни. В мире впервые трактовка понятия ЧСВ встречается в трудах Карлоса Кастанеды. В третьей книге «Путешествие в Икстлан» и других трудах говорится о том, что ЧСВ не является врожденным.
В третьей книге «Путешествие в Икстлан» и других трудах говорится о том, что ЧСВ не является врожденным.
На протяжении жизни, особенно в подростковом возрасте, оно приобретается, как ответ на взаимодействие с внешним миром. Под воздействием чувства важности у человека меняются приоритеты, остаются только те из них, которые согласованы с ЧСВ. Под воздействием ложных установок человек способен даже на неразумные вредящие жизни поступки.
Состояние характеризуется, как деструктивное, оно порождает действия, направленные только на поддержания собственной значимости. Потеря ЧСВ — одно из условий для принятия ответственности за свои поступки. По словам Кастанеды, так появляется состояние воина. Особое состояние характеризуется, как рассредоточение общего внимания вместе с состоянием внутренней мобилизации.
ЧСВ это расшифровка в молодежной среде означает — чувство собственной важности.Ярким примером такого чувства может служить момент погружения в интересную работу. Внутренние силы достигают наивысшей точки эффективности, действия продуктивны, человек, занимающийся любимым делом, в такие минуты счастлив.
Примеры использования в речи
У людей старшего поколения, воспитанного пионерией и комсомолом, подобное поведение встречается крайне редко.
Общаясь с молодежью, можно услышать следующие язвительные выражения:
- — Похоже, что ЧСВ ты имеешь завышенное!;
- — У тебя ЧСВ просто зашкаливает, нужно быть попроще;
- — Перестань повышать свое ЧСВ за счет унижения других;
- — У тебя заниженное ЧСВ. Ты явно себя не ценишь.
Даже если среди знакомых не принято употребление выражения, его значение нужно знать, чтобы не попадать в глупое положение. Любое не правильно понятое слово – повод для нарушения коммуникации. Разговор теряет смысл, нужные вопросы не решаются.
ЧСВ — это хорошо или плохо?
Иногда ЧСВ путают с чувством собственного достоинства. Личность с развитым чувством собственного достоинства приятна в общении и дружелюбна по отношению к окружающим. Присутствует уважение как к собственной персоне, так и к другим. Когда хотят задеть чувство собственного достоинства, специально задевают, чтобы обидеть.
Когда хотят задеть чувство собственного достоинства, специально задевают, чтобы обидеть.
Критика собственной важности направлена исключительно на эту черту. Избавившись от ЧСВ, человек становится по-настоящему неуязвимым. Для беспочвенных обвинений больше нет повода. А беспочвенная критика больше не отнимает силы на оправдание и доказательство собственного всестороннего превосходства.
Чем опасно ЧСВ
Человек, подверженный этому пороку, считает себя лучшим и самым сведущим, без веских на то оснований, а это прямой путь к деградации. Занимая подобную позицию, создается зашоренность восприятия действительности. Не принимая во внимание чужую точку зрения, велика вероятность пропустить рациональное зерно, как следствие, в тяжелых случаях может наступить отсутствие развития.
Замыкаясь в себе, не принимая и не прислушиваясь к чужой точке зрения, наоборот, постоянно всех критикуя и оспаривая чужое мнение, есть опасность растерять настоящих друзей и в итоге остаться в одиночестве. Такое поведение вызывает у других в лучшем случае недоумение, в худшем – отторжение.
Такое поведение вызывает у других в лучшем случае недоумение, в худшем – отторжение.
Таким образом, негативные стороны влияния ЧСВ можно свести к нескольким пунктам:
- ухудшение отношений с родными, друзьями и коллегами;
- прогрессирующее сужение круга общения;
- выделение из общего ряда только тех событий и фактов, которые связаны с его собственной персоной;
- неприятие чужой точки зрения, как следствие остановка в саморазвитии.
Современный человек может жить и развиваться только в общении с другими.
Завышенное ЧСВ
Определить, что у кого-либо из окружения есть такая нехорошая черта, можно по следующим признакам:
- индивидуум навязывает всем свою точку зрения, выступая при этом в роли наставника;
- слишком много вопросов решается спором, агрессивным отстаиванием своей точки зрения;
- привлечение всеобщего внимания к собственной персоне, стремление всегда быть в центре внимания, порой любой ценой;
- мстительность, как яркая черта характера.
 Стремление мстить за критику, даже если она была справедливой;
Стремление мстить за критику, даже если она была справедливой; - пристальное внимание к чужим ошибкам, стремление возвыситься, унижая других.
 Стремление мстить за критику, даже если она была справедливой;
Стремление мстить за критику, даже если она была справедливой;К счастью, в реальной жизни таких персонажей не так много, чего не скажешь о виртуале. В среде, где легко спрятаться за ярким ником, как через увеличительную призму видны все худшие наклонности. Самое большое число людей с завышенным чувством величия наблюдается в среде известных медиаперсон.
Они настолько увлекаются той ролью, которую играют на публике, что переносят подобный стиль общения и в обычную жизнь. В то же время есть случаи, когда доказывать собственную значимость необходимо по той роли, которую человек занимает в обществе.
Завышенным ЧСВ не является для следующих категорий:
- люди опасных профессий, например, военные живут по своим «законам». Если человек с более высоким званием требует беспрекословного подчинения, его поведение продиктовано соблюдением субординации и порядка, на которых и держится дисциплина;
- шарлатан, продающий липовое лекарство, будет раздувать свое ЧСВ до бесконечности. Он делает это только для того, чтобы легковерные покупатели не сомневались в аргументированности его слов;
- жесткость критики экспертов в чем-либо основана на доскональном знании предмета спора и с личными характеристиками не имеет ничего общего.
 Он делает это только для того, чтобы легковерные покупатели не сомневались в аргументированности его слов;
Он делает это только для того, чтобы легковерные покупатели не сомневались в аргументированности его слов;Та роль, которую люди играют в обществе, может характеризовать стиль поведения, сходный с ЧСВ. Но к характеру это не имеет отношения, только если люди умеют вне своей профессиональной деятельности абстрагироваться от работы.
Заниженное ЧСВ
В некоторой доле это чувство должно присутствовать у каждого человека. Заниженное ЧСВ так же опасно, так как во всем важно поддерживать баланс. Нужно прислушиваться и доверять чужому мнению, но и не поступаться собственными принципами. В виртуале важно не реагировать на заведомо пустые в плане разумного продолжения споры и вопросы.
Подростковое ЧСВ
ЧСВ (расшифровка в молодежной среде иногда звучит, как Чувство собственного величия) по отношению к подростку чаще всего используется справедливо. И на то есть свои причины. Подростку важно завоевать внимание окружающих. В силу малого опыта методы завоевания всеобщего одобрения могут быть не адекватными.
И на то есть свои причины. Подростку важно завоевать внимание окружающих. В силу малого опыта методы завоевания всеобщего одобрения могут быть не адекватными.
Девушки любят говорить о себе и находятся в постоянном поиске молодого человека, который оценит их уникальность. Парни, кроме разговоров о собственной персоне, любят выстраивать теории и делиться собственным мнением. В период взросления молодые люди очень болезненно реагируют на критику. Их главной целью может быть самоутверждение в обществе себе подобных.
Им свойственна заносчивость, неуживчивость, неумение прислушиваться к чужой точке зрения. Стремясь любой ценой занять положение лидера в компании, такие молодые люди буквально «идут по головам». К ним чаще всего применяется язвительное замечание, собственно в этой среде оно и появилось и чаще всего используется.
ЧСВ в интернете
В реальной жизни есть больше поводов задуматься о том, какая реакция будет в ответ на сказанные слова и поступки. В виртуале появляется ложное чувство безнаказанности. Можно легко сменить ник, свой IP адрес.
Можно легко сменить ник, свой IP адрес.
Проявляются худшие черты характера, порой в гипертрофированном виде. Иногда, когда есть возможность познакомиться с человеком в реальной жизни, возникает чувство удивления, насколько это приятный в общении человек.
ЧСВ в ВК
ЧСВ (расшифровка в молодежной среде знакома большинству обладателей акккаунтов в ВК) как черта характера высмеивается в комментариях и многочисленных картинках. Реакция на несправедливое замечание только подливает масла в огонь. Если есть уверенность, что замечание беспочвенное, лучше не реагировать.
Бурная реакция дает повод высмеивать и дальше, так как оправдания – один из признаков желания казаться безупречным и лучше других.
Картинки с тематическими рисунками гуляют по сети. Отвечая на одну, есть шанс получить еще несколько вдогонку. Есть возможность отгородиться от шутника настройками профиля, но это тоже не выход. Вместе с ним можно лишиться и нужных знакомых. Самая лучшая тактика – отвечать предельно дружелюбно и не поддаваться на провокации.
ЧСВ в играх
Большинство интерактивных игр основано на командной поддержке. Игрок, с завышенным ЧСВ не считается с интересами других игроков, команда в итоге проигрывает от его же действий.
А в ответ на справедливые обвинения обычно звучит что-то вроде: «Я сделал все как надо, а другие действовали неправильно, недостаточно быстро, продуманно и так далее». Естественно, что в ответ зарвавшемуся игроку «прилетит» обвинение в ЧСВ.
Афоризмы и фразы людей с ЧСВ
Люди с повышенным ЧСВ часто пишут следующие фразы:
- Я лучший в том-то.
- Мои действия самые правильные, самые результативные.
- Я делаю так, как считаю нужным, мнение остальных неправильное.
- Ну и что, что кто-то больше написал (работал, думал), я-то лучше!
Отсутствие авторитетов и внимания к остальным сразу обозначают подобных участников.
Как реагировать на замечание?
Любая критика, в том числе и обвинение в «распухшем» ЧСВ, больно ранит и вызывает неприятные эмоции.
Есть 3 неправильных способа ответа на критику:
- оправдываясь, человек занимает униженную позицию. К сожалению, это самый распространенный способ реагирования, примерно 60%. Как только человек начинает оправдываться, у оппонента возникает впечатление, что он уже признает свою вину;
- контратака, обвинение собеседника в том же, в чем он обвиняет других. Такая реакция встречается в 20% случаев. Этот вид реагирования позволяет сохранить лицо, но безвозвратно портит отношения;
- молчание в ответ позволяет сохранить лицо и не испортить окончательно отношения. Такая реакция встречается в 10% случаев. Единственный недостаток, который сводит «на нет» все преимущества такого способа реагирования состоит в том, что человек, смолчавший в подобной ситуации, особенно если обвинения безосновательны, наносит большой вред своему здоровью. Человеку необходима разрядка после необоснованной критики, любое активное действие, хотя бы разорванный лист бумаги. Главное — не срываться на живых людях.

Проще всего игнорировать, не принимая замечания близко к сердцу, но это не конструктивный путь и прямая возможность потерять отношения с человеком.
Правильная реакция на язвительное замечание зависит от справедливости обвинения:
- в случае полной уверенности в необоснованности обвинения, следует задать встречный вопрос, на чем именно базируется данное обвинение. Тон вопроса должен быть предельно доброжелательным, нельзя ехидничать и ни в коем случае оскорблять в ответ. Такая форма общения может стать конструктивным продолжением разговора, конечно, если оппонент настроен на продолжение разговора;
- если есть вероятность, что своими действиями человек, подвергшийся подобной критике, мог сам ее спровоцировать, нужно частично согласиться, стараясь свести разговор к безобидной шутке, например, похвалив наблюдательность оппонента;
- в случае если человек явно не настроен на доброжелательное продолжение разговора, ответные фразы не должны содержать ничего лишнего. Нельзя давать оппоненту почвы для продолжения критики, должна быть только дружелюбная ответная реакция. Фразы должны быть короткими не более 5 слов, не должно быть выражений типа «Да…, но», потому что подобная конструкция воспринимается как отрицание и дает почву для продолжения конфликта.
 Нельзя давать оппоненту почвы для продолжения критики, должна быть только дружелюбная ответная реакция. Фразы должны быть короткими не более 5 слов, не должно быть выражений типа «Да…, но», потому что подобная конструкция воспринимается как отрицание и дает почву для продолжения конфликта.
Нельзя давать оппоненту почвы для продолжения критики, должна быть только дружелюбная ответная реакция. Фразы должны быть короткими не более 5 слов, не должно быть выражений типа «Да…, но», потому что подобная конструкция воспринимается как отрицание и дает почву для продолжения конфликта.У любого конфликта есть положительная конструктивная сторона. Есть возможность взглянуть на себя чужими глазами. В случае если велика вероятность, что обвинения не беспочвенны, есть смысл пересмотреть свои приоритеты. Не так часто, но бывает, что человек, указавший на недостаток, желает только добра.
Как обуздать чувство собственной важности
Увидеть себя «со стороны» всегда полезно, а реагировать на конструктивную критику — значит идти по пути улучшения.
Побороть плохую черту можно, совершив несколько последовательных шагов:
- чистый лист нужно поделить на 2 равные части: справа пишутся положительные черты, слева – отрицательные;
- нужно подвергнуть анализу последние конфликтные ситуации с точки зрения справедливости занятой точки зрения;
- на другом чистом листе нужно написать имена тех, кто был незаслуженно обижен словами и действиями тестируемого;
- необходимо сделать выводы и сформулировать новую линию поведения, без ущемления чужого достоинства;
- для мотивации можно заключить с собой пари на выполнение поставленной задачи по изменению собственной линии поведения.

Изменить себя — сложная задача, но при правильном плане действий она по силам любому. Другая сторона медали, когда у человека заниженное ЧСВ.
Для достижения гармонии подойдут следующие шаги:
- в спорах, где человек заведомо более сведущий, нужно отстаивать свою точку зрения, это пойдет на пользу всем. Человек с заниженным ЧСВ приобретает так недостающую ему уверенность в себе и собственных знаниях. А остальные участники спора получают ценную информацию от близкого им человека;
- нужно останавливать явно зарвавшихся спорщиков. Проглатывание обиды пагубно сказывается на здоровье и на репутации неуверенного в себе человека;
- не стыдно подкреплять свои слова, ссылаясь на проверенные источники, если нет уверенности, что именно к словам говорящего прислушаются. Можно аргументировать совпадающей точкой зрения с уважаемыми общими знакомыми;
- похвала — хороший повод исправить ситуацию. Не нужно прятаться, когда хвалит начальник. Скромность хороша в меру, как и остальные черты;
- необходимо научиться игнорировать язвительные замечания, особенно когда они адресованы только с целью вывести из равновесия.
 Скромность хороша в меру, как и остальные черты;
Скромность хороша в меру, как и остальные черты;В момент, когда ЧСВ приходит в норму, больше не вредит окружающим и не загоняет в жесткие рамки неуверенного в себе человека, высвобождается масса энергии. Есть хорошая предпосылка для рывка в саморазвитии и достижении поставленных целей.
Как ни парадоксально, но именно люди, увлекающиеся литературой о личностном росте, особенно в начале увлечения, страдают от увеличенной важности.
Нужно изначально быть развитой личностью, чтобы подобная информация легла на благодатную почву. Книги Карлоса Кастанеда «Путешествие в Икстлан», «Отдельная реальность» и «Сказки о силе» будут интересны людям исследующим духовные практики. Можно равнодушно относиться к большинству мемов в молодежной среде.
Они возникли на благодатной почве виртуального общения, обычно в виртуальном мире они и остаются. ЧСВ неотделимо от реальной жизни, его расшифровка в разных группах общества имеет общие черты. Правильная установка самоопределения в обществе способна придать новый толчок к развитию.
ЧСВ неотделимо от реальной жизни, его расшифровка в разных группах общества имеет общие черты. Правильная установка самоопределения в обществе способна придать новый толчок к развитию.
В то же время, необоснованно завышенное ЧСВ, как психологически стойкая особенность характера человека, по мнению психотерапевтов, становится проблемой не только и не столько для окружающих, сколько для него самого.
Кроме того, что портятся отношения с важными в жизни людьми, эта черта в своем гипертрофированном виде отнимает много сил и энергии, которые с пользой можно было бы направить в другое более продуктивное русло.
Оформление статьи: Анна Винницкая
Видео о расшифровке ЧСВ
Как распознать ЧСВ:
биография, фото богини ЧСВ, номер Маши Сорокиной
| Полное имя | Маша Сорокина |
| Род деятельности | блогер |
| Дата рождения | 04. 02.2005 02.2005 |
| Знак зодиака | Водолей |
| Рост/Вес | неизвестно |
| Семейное положение | не замужем |
| Инстаграм | Ссылка |
| Youtube | Ссылка |
| Википедия | — |
Мерлин (Merelin) – маленькая лайкерша из северной столицы России, стремительно набирающая популярность в интернете. Величает себя богиней, поэтому некоторые подписчики считают ее высокомерной.
Биография Мерлины из Лайка. Какое настоящее имя и сколько лет?
Маша Сорокина (настоящее имя девочки) родилась 4 февраля 2005 года. У Марии есть сестра Настя, она также занимается блогерством, но не столь успешна как Merelin. Обе сестрички посещают Санкт-Петербургский государственный «Театр детского балета». Известно, что Маша также училась на подготовительном отделении в Академии русского балета имени А. Я. Вагановой. В 2018 году принимала участие в Международном фестивале-конкурсе хореографического мастерства «Эталон».
Я. Вагановой. В 2018 году принимала участие в Международном фестивале-конкурсе хореографического мастерства «Эталон».
Известность получила благодаря топовому приложению Лайк, огромный всплеск популярности приходится на январь 2020 года. Такого фантастического роста подписчиков не наблюдалось ни у одного лайкера и сейчас Merelin является одним из лидеров русскоязычного сегмента. В ее видеороликах можно увидеть сестру и маму.
Маша с сестричкой НастейОна является солисткой балета в своем театре, поэтому хорошо умеет играть на публику и прекрасно двигается. Наверное, эти качества привели к стремительному успеху. Но у Маши немало хейтеров, называющих ее тщеславной, эгоцентричной и самовлюбленной девочкой с высоким ЧСВ (чувством собственной важности). Такие люди просто воспринимают все всерьез, но девочка удачно выбрала образ. В Лайке тысячи блогеров, выполняющих однотипные действия и поэтому не запоминающихся аудитории. Мария (или человек, который ей посоветовал) сделала правильную ставку. Merlin говорит, что она ярче других, что она – Богиня (или королева). Она строит гримасы, надменно называет подписчиков нищебродами, рассказывает, что она богатая, живет в крутом доме. Но это она делает по сценарию, создавая шуточный образ. Фактически Маша высмеивает золотую молодежь («мажоров»), но многие подростки и дети принимают шутки близко к сердцу и начинают хейтить ее. Однако даже отрицательный пиар положительно сказывается на ее рейтинге, и девчонка стала прилично зарабатывать, у нее уже присутствует собственный менеджер. Она принимает участие в сходках и других лайкерских мероприятиях.
Merlin говорит, что она ярче других, что она – Богиня (или королева). Она строит гримасы, надменно называет подписчиков нищебродами, рассказывает, что она богатая, живет в крутом доме. Но это она делает по сценарию, создавая шуточный образ. Фактически Маша высмеивает золотую молодежь («мажоров»), но многие подростки и дети принимают шутки близко к сердцу и начинают хейтить ее. Однако даже отрицательный пиар положительно сказывается на ее рейтинге, и девчонка стала прилично зарабатывать, у нее уже присутствует собственный менеджер. Она принимает участие в сходках и других лайкерских мероприятиях.
Известен ли номер Мерлин?
Во вконтакте есть группы, где пользователи (наверное, знакомые) делятся номерами известных лайкеров, например, Лисички Элины и Миланы Некрасовой. Номер Merelin неизвестен, найти ее в приложении можно по никнейму, указанному ниже.
Социальные сети Merelin из Likee
Мерлин есть в следующих соцсетях (данные на начало 2020 года):
- Инстаграм (ник mariyasor_rg), 117 тысяч подписчиков;
- Ютуб (MERELIN MikeRahe), 37 тыс. фолловеров;
- Собственно Like (mera774nf), 5,4 миллиона подписанных пользователей.
 фолловеров;
фолловеров;Из-за мегаизвестности лайкерши многие юзеры начали создавать страницы с аналогичным никнеймом, поэтому если вы введёте «Merilin» в поисковую строку, вы не сразу обнаружите подлинную страничку. Многие из клонов набирают тысячи и десятки тысяч подписчиков, но до пятимилионной аудитории Маши им, безусловно, далеко.
В Instagram девочка постит преимущественно видео, но фото тоже присутствуют. Особое внимание фанов привлекают совместные фотографии с другими знаменитостями Лайка, такими как Настя Кош и Макар Карелин. Фоток с родителями Мерелин не публикует, но однажды запостила снимок с бабушкой, похваставшись удачным походом в лес за грибами. Инстаграм-аккаунт существует у нее с 2016 года, поэтому если пролистать его далеко вниз, можно увидеть фото маленькой Мерлин (в детстве). В основном лента «ранней Маши» была посвящена балету, она демонстрировала великолепную растяжку и превосходное мастерство на сцене.
Дорогие пользователи, если вам известен ВК героини этой статьи, сообщите, пожалуйста в комментариях.
Лайкер Маша Сорокина сейчас
Девочка активно общается с подписчиками в Лайке и Инстаграме. В инсте они присылают ей рисунки/картинки с ее изображением и справедливо отмечают, что здесь она «не такая ЧСВ», что только подтверждает вымышленность скандального образа в Like.
Недавно Мария с сестричкой Анастасией отметили пятнадцатилетие (некоторые порталы пишут, что возраст блогерши 12 лет, но она сама написала, что ей 15). Они получили массу поздравлений от поклонников, а другие маленькие блогеры записали в честь праздника видеоролики.
Источник изображений: Инстаграм Маши Сорокиной из лайка.
Merelin – феномен Лайка со взрывным ростом популярности
3.7 (73.33%) 15 votesлабораторий злого гения | Учебное пособие по FastLED RGB и HSV
Фон
Я готовлюсь к участию в Maker Faire Kansas City 2017 и пытаюсь создать интерактивный образовательный проект, с которым люди могли бы поиграть.
Имея под рукой то, что у меня было, я быстро собрал пару макетов, каждая с Teensy, несколькими светодиодами RGB и тремя потенциометрами (используя отличный прочный адаптер потенциометра для макетов Пола Стоффрегена).
Хотя это и сработает, но он не очень прочный.Мне нужно что-то, с чем многие люди могут возиться и играть все выходные, не беспокоясь о замене отключенных проводов.
Итак, я решил спроектировать печатную плату, которую я смогу собрать заранее и, надеюсь, будет достаточно прочной, чтобы прослужить все выходные. У него также будет бонус в виде учебного проекта, который я могу использовать на занятиях, семинарах и делиться онлайн.
Содержание
Требования к проекту
- Образовательный: Я хочу, чтобы люди учились чему-то, пока возятся с этим.
- Долговечность: Потребуется много злоупотреблений.
- Открыто: Это может противоречить долговечности, но я действительно не хочу, чтобы это было спрятано в корпусе. Я хочу, чтобы кишки были видны.
- Открытый исходный код и оборудование: Я хотел бы поделиться этим как можно более свободно и широко.
- Недорого: В идеале 30 долларов или меньше, чтобы люди могли позволить себе их построить.
- Качество: Может противоречить недорогим ценам, но нет смысла строить их дешево, если они недолговечны.
- Доступно для детей: Легко собрать, но настолько легко, что ничему не научишься, собирая его. Все компоненты должны быть сквозными. Не требуется пайки для поверхностного монтажа, печи оплавления и т. Д.
 Я хочу, чтобы кишки были видны.
Я хочу, чтобы кишки были видны.Помня об этих целях, я создал эту печатную плату.
Печатная плата
Вот файлы Eagle (схема и плата)
Характеристики
Переключатель переключает потенциометры с управления RGB на HSV.Печатная плата включает в себя конденсатор и резистор NeoPixel «передовой опыт», а также контакты для подключения большего количества светодиодов (полоса, кольцо и т. Д.). Тактильная кнопка мгновенного действия может использоваться для переключения режимов, от учебного курса до шаблонов, анимации и т. Д.
Д.). Тактильная кнопка мгновенного действия может использоваться для переключения режимов, от учебного курса до шаблонов, анимации и т. Д.
Монтажные отверстия под Teensy LC позволяют устанавливать печатные платы на плате, чтобы они не упали со стола или не «ушли».
Список деталей
Компоненты
Светодиоды
Я решил использовать NeoPixel Stick 8x.Они немного дешевле и включают больше светодиодов, чем NeoPixels для макетных плат.
Микропроцессор
Стремясь минимизировать стоимость деталей при сохранении простоты сборки и использования, я сначала подумал об использовании 5-вольтового брелка. Они недорогие, но все же высокого качества и не требуют переключателя уровня. У них просто не хватает входов, поэтому я переключился на 5V Pro Trinket. Затем я вспомнил Teensy LC, который стоит меньше, чем на 2 доллара, имеет цифровой выходной вывод 5 В, более быстрый процессор, больше ОЗУ, флэш-памяти и т. Д.Его также можно заказать в OSH Park вместе с печатной платой. Несколько человек также предупредили меня, что процесс загрузки брелка может быть немного утомительным.
Д.Его также можно заказать в OSH Park вместе с печатной платой. Несколько человек также предупредили меня, что процесс загрузки брелка может быть немного утомительным.
Пайка
Примечание : Тщательно проверьте положение, выравнивание и ориентацию каждого компонента перед пайкой!
Если вы новичок в пайке, я настоятельно рекомендую покупать Teensy LC с уже припаянными контактами. Это сэкономит вам время и силы.
Я также рекомендую прочитать хорошее руководство по пайке, например, от Adafruit и SparkFun.
Teensy LC
Осторожно припаяйте контакты заголовка 0,1 дюйма (2,54 мм) к Teensy LC. Проще всего вставить контакты заголовка в макет и припаять оттуда Teensy, как показано в этом руководстве от SparkFun.
Вы также можете использовать печатную плату, чтобы выровнять и припаять контакты после того, как вы добавили женские разъемы, если у вас нет макетной платы.
Вам также нужно будет припаять 5 выводов на конце Teensy. По крайней мере, вы должны припаять один контакт разъема к отверстию для контакта 17 на конце. Это логический выходной контакт 5 В, используемый для управления NeoPixels.
По крайней мере, вы должны припаять один контакт разъема к отверстию для контакта 17 на конце. Это логический выходной контакт 5 В, используемый для управления NeoPixels.
Печатная плата
Я предпочитаю добавлять компоненты к печатной плате от самых коротких до самых высоких. Я считаю, что таким способом легче всего удерживать печатную плату устойчиво при пайке. Если у вас есть третья рука, это тоже может помочь.
Тактильная кнопка
Если вы используете тактильную кнопку, которая короче резисторов, сначала вставьте и припаяйте ее.
Резисторы
Согните одну ножку каждого резистора на 180 градусов и вставьте их в соответствующие места на печатной плате. Убедитесь, что вы поместили их в правильные места, как указано на печатной плате. Припаяйте их на место.
Переключатель
Если у вашего переключателя есть маленькие металлические выступы на конце (например, от Adafruit), вам может потребоваться обрезать их или согнуть, чтобы переключатель встал заподлицо на печатной плате. Припаяйте переключатель к плате, как указано.Ориентация переключателя не имеет значения.
Припаяйте переключатель к плате, как указано.Ориентация переключателя не имеет значения.
Головки с внутренней резьбой
Если у вас есть макетная плата и несколько удлиненных штыревых контактов разъема, вы можете использовать их для выравнивания и удержания разъемов с внутренней резьбой во время пайки.
Вы также можете вставить Teensy LC в гнездовые разъемы, а затем вставить женские разъемы в печатную плату, чтобы убедиться, что они правильно выровнены.
Вставьте, выровняйте и припаяйте гнезда разъемов. Перед пайкой убедитесь, что они правильно выровнены!
Тактильная кнопка
Если вы используете тактильную кнопку, которая выше резисторов, вставьте и припаяйте ее сейчас.
Конденсатор
Вставить и припаять конденсатор. Примечание: Убедитесь, что отрицательный вывод, помеченный знаком минус (-), вставлен в отмеченное отрицательное отверстие на печатной плате!
Потенциометры
Вставьте и припаяйте потенциометры.
NeoPixels
Используйте третью руку, прищепку, канцелярскую скрепку и т. Д., Чтобы удерживать короткую сторону штырей под прямым углом на стороне «DIN» полосы NeoPixel во время пайки.Более длинные ножки штифтов с черным пластиковым изолятором должны выступать под прямым углом к стержню NeoPixel.
Сборка
Вставьте Teensy LC в разъемы на печатной плате. Убедитесь, что он правильно сориентирован, а порт USB находится на краю печатной платы.
Вставьте полоску NeoPixel в гнезда. Еще раз проверьте выравнивание, сравнив метки на полосе с метками на печатной плате.
Ручки для потенциометров хороши, но не обязательны.Я купил несколько дешевых ручек на Amazon, но мне пришлось обрезать их кусачками и немного просверлить центральное отверстие, чтобы они подошли.
Вы также можете нанести немного красной, зеленой и синей краски или лака для ногтей на верхнюю часть штырей потенциометра.
Если у вас есть мультиметр, который может измерять сопротивление, рекомендуется после сборки убедиться в отсутствии коротких замыканий между питанием и землей на плате. Вы должны измерить бесконечное (или, по крайней мере, очень высокое) сопротивление между выводами 5V и GND на Teensy LC, полосе NeoPixel, разъеме и т. Д.
Вы должны измерить бесконечное (или, по крайней мере, очень высокое) сопротивление между выводами 5V и GND на Teensy LC, полосе NeoPixel, разъеме и т. Д.
После двойной проверки правильности совмещения всех компонентов вы можете подключить Teensy LC к компьютеру с помощью кабеля Micro USB.
Настройка программного обеспечения
Ардуино IDE
Загрузите и установите или извлеките (Arduino IDE] (https://www.arduino.cc/en/Main/Software).
Teensyduino
Чтобы скомпилировать скетч для Teensy, вам понадобится надстройка для Arduino IDE под названием Teensyduino. Скачайте и установите его.
Вам будет предложено установить дополнительные библиотеки во время установки Teensyduino. Не устанавливайте все библиотеки! Щелкните «Нет».
Библиотека FastLED
Если вы не знакомы с ними, руководство Adafruit по библиотекам Arduino расскажет вам все, что вы когда-либо хотели знать о библиотеках, включая более подробные инструкции по установке.
Загрузите последнюю версию библиотеки FastLED, затем распакуйте файл и переименуйте папку в «FastLED» (без кавычек).Скопируйте файл в папку Arduino / Libraries.
Или используйте git для клонирования репозитория FastLED:
cd ~ / Documents / Arduino / библиотеки
git clone https://github.com/FastLED/FastLED.git
В любом случае вам, вероятно, потребуется перезапустить Arduino, чтобы библиотека загрузилась и появилась.
Код
Я создал простой эскиз Arduino, чтобы показать, как работают RGB и HSV для создания разных цветов. Я также включил радужный узор, который отображается при удерживании нажатой тактильной кнопки мгновенного действия.
Скопируйте и вставьте код в новый скетч Arduino:
Компиляция
Настроить Arduino
- В среде Arduino IDE щелкните меню Tools , затем Board , затем щелкните Teensy LC .
- Снова щелкните меню Tools , затем Port , а затем порт, к которому подключен Teensy LC.
- Нажмите кнопку Проверить (флажок) на панели инструментов.
- Он должен закончиться и сказать Готово, в строке состояния в нижней части окна.
- Теперь нажмите кнопку Загрузить (стрелка вправо) на панели инструментов.

Более подробная информация в ближайшее время…
| enum | HTMLColorCode { AliceBlue = 0xF0F8FF, Аметист = 0x9966CC, AntiqueWhite = 0xFAEBD7, Аква = 0x00FFFF, Аквамарин = 0x7FFFD4, Лазурный = 0xF0FFFF, Бежевый = 0xF5F5DC, Биск = 0xFFE4C4, Черный = 0x000000, Бланшированный миндаль = 0xFFEBCD, Синий = 0x0000FF, Синий-фиолетовый = 0x8A2BE2, Коричневый = 0xA52A2A, BurlyWood = 0xDEB887, CadetBlue = 0x5F9EA0, Шартрез = 0x7FFF00, Шоколад = 0xD2691E, Коралл = 0xFF7F50, Василек синий = 0x6495ED, Cornsilk = 0xFFF8DC, Малиновый = 0xDC143C, Голубой = 0x00FFFF, Темно-синий = 0x00008B, DarkCyan = 0x008B8B, DarkGoldenrod = 0xB8860B, темно-серый = 0xA9A9A9, темно-серый = 0xA9A9A9, Темно-зеленый = 0x006400, Темный хаки = 0xBDB76B, Темно-пурпурный = 0x8B008B, DarkOliveGreen = 0x556B2F, Темно-оранжевый = 0xFF8C00, DarkOrchid = 0x9932CC, Темно-красный = 0x8B0000, Темный лосось = 0xE9967A, DarkSeaGreen = 0x8FBC8F, DarkSlateBlue = 0x483D8B, DarkSlateGray = 0x2F4F4F, DarkSlateGrey = 0x2F4F4F, Темно-бирюзовый = 0x00CED1, Темно-фиолетовый = 0x9400D3, DeepPink = 0xFF1493, DeepSkyBlue = 0x00BFFF, DimGray = 0x696969, DimGrey = 0x696969, DodgerBlue = 0x1E90FF, FireBrick = 0xB22222, Цветочно-белый = 0xFFFAF0, Лесозеленый = 0x228B22, Фуксия = 0xFF00FF, Гейнсборо = 0xDCDCDC, GhostWhite = 0xF8F8FF, Золото = 0xFFD700, Золотарник = 0xDAA520, Серый = 0x808080, Серый = 0x808080, Зеленый = 0x008000, зеленый желтый = 0xADFF2F, Ханидью = 0xF0FFF0, HotPink = 0xFF69B4, Индийский красный = 0xCD5C5C, Индиго = 0x4B0082, слоновая кость = 0xFFFFF0, хаки = 0xF0E68C, Лаванда = 0xE6E6FA, LavenderBlush = 0xFFF0F5, LawnGreen = 0x7CFC00, Лимонный шифон = 0xFFFACD, Голубой = 0xADD8E6, LightCoral = 0xF08080, LightCyan = 0xE0FFFF, LightGoldenrodYellow = 0xFAFAD2, Светло-зеленый = 0x90EE90, Светло-серый = 0xD3D3D3, LightPink = 0xFFB6C1, светлый лосось = 0xFFA07A, LightSeaGreen = 0x20B2AA, LightSkyBlue = 0x87CEFA, LightSlateGray = 0x778899, LightSlateGrey = 0x778899, LightSteelBlue = 0xB0C4DE, Светло-желтый = 0xFFFFE0, Лайм = 0x00FF00, зеленовато-желтый = 0x32CD32, Белье = 0xFAF0E6, пурпурный = 0xFF00FF, Бордовый = 0x800000, Средний Аквамарин = 0x66CDAA, MediumBlue = 0x0000CD, Средняя орхидея = 0xBA55D3, средний фиолетовый = 0x9370DB, MediumSeaGreen = 0x3CB371, MediumSlateBlue = 0x7B68EE, MediumSpringGreen = 0x00FA9A, Средний Бирюзовый = 0x48D1CC, средний фиолетовый красный = 0xC71585, MidnightBlue = 0x191970, MintCream = 0xF5FFFA, MistyRose = 0xFFE4E1, Мокасин = 0xFFE4B5, Навахо Белый = 0xFFDEAD, ВМС = 0x000080, OldLace = 0xFDF5E6, Оливка = 0x808000, OliveDrab = 0x6B8E23, Оранжевый = 0xFFA500, Оранжевый Красный = 0xFF4500, Орхидея = 0xDA70D6, PaleGoldenrod = 0xEEE8AA, PaleGreen = 0x98FB98, PaleTurquoise = 0xAFEEEE, PaleVioletRed = 0xDB7093, Кнут папайи = 0xFFEFD5, PeachPuff = 0xFFDAB9, Перу = 0xCD853F, Розовый = 0xFFC0CB, Плед = 0xCC5533, Слива = 0xDDA0DD, PowderBlue = 0xB0E0E6, Фиолетовый = 0x800080, Красный = 0xFF0000, розово-коричневый = 0xBC8F8F, RoyalBlue = 0x4169E1, Седло Коричневый = 0x8B4513, Лосось = 0xFA8072, SandyBrown = 0xF4A460, SeaGreen = 0x2E8B57, Морская ракушка = 0xFFF5EE, Сиенна = 0xA0522D, Серебро = 0xC0C0C0, SkyBlue = 0x87CEEB, SlateBlue = 0x6A5ACD, серый серый = 0x708090, SlateGrey = 0x708090, Снег = 0xFFFAFA, SpringGreen = 0x00FF7F, SteelBlue = 0x4682B4, Желто-коричневый = 0xD2B48C, бирюзовый = 0x008080, Чертополох = 0xD8BFD8, Помидор = 0xFF6347, Бирюза = 0x40E0D0, Фиолетовый = 0xEE82EE, Пшеница = 0xF5DEB3, Белый = 0xFFFFFF, Белый дым = 0xF5F5F5, Желтый = 0xFFFF00, желто-зеленый = 0x9ACD32, FairyLight = 0xFFE42D, FairyLightNCC = 0xFF9D2A } |
Клеточное программирование HSV-2 делает возможным продуктивную ВИЧ-инфекцию в дендритных клетках
Front Immunol. 2019; 10: 2889.
2019; 10: 2889.
Элиза Криши
1 Отдел молекулярной вирусологии, Департамент клинической и экспериментальной медицины, Университет Линчёпинга, Линчёпинг, Швеция
Сесилия 1 Отделение молекулярной вирусологии, Отдел клинической и экспериментальной медицины, Университет Линчёпинга, Линчёпинг, ШвецияRada Ellegård
1 Отделение молекулярной вирусологии, Отдел клинической и экспериментальной медицины, Университет Линчёпинга, Линчёпинг, Швеция
Мохаммад Халид
1 Отдел молекулярной вирусологии, Департамент клинической и экспериментальной медицины, Университет Линчёпинга, Линчёпинг, Швеция
906 13 2 Кафедра фармацевтики, Фармацевтический колледж, Университет Короля Халида, Абха, Саудовская Аравия
Джулия Хеллблом
1 Отделение молекулярной вирусологии, Кафедра клинической и экспериментальной медицины, Университет Линчёпинга, Линчёпинг, Швеция
Казуки Окуяма
3 Отделение экспериментальной гематологии, Отдел клинической и экспериментальной медицины, Университет Линчёпинга, Линчёпинг, Швеция
Прадёт Бхаттачарья
1 Отделение молекулярной вирусологии, Отдел клинической и экспериментальной медицины, Университет Линчёпинга, Линчёпинг, Швеция
Sofia Nyström
1 Отделение молекулярной вирусологии, отделение клинической и экспериментальной медицины, Университет Линчёпинга, Линчёпинг, Швеция
Esaki M.
 Шанкар
Шанкар4 Отделение инфекционной биологии и медицинской микробиологии, Департамент наук о жизни, Школа наук о жизни, Центральный университет Тамил Наду, Тируварур, Индия
Кристина Эрикссон
5 Департамент исследований ревматологии и воспаления, Университет of Gothenburg, Gothenburg, Sweden
Marie Larsson
1 Отделение молекулярной вирусологии, Отдел клинической и экспериментальной медицины, Университет Линчёпинга, Линчёпинг, Швеция
1 Отделение молекулярной вирусологии, Отдел клинической и экспериментальной медицины, Университет Линчёпинга, Линчёпинг, Швеция
2 Кафедра фармацевтики, Фармацевтический колледж, Университет Короля Халида, Абха, Саудовская Аравия
3 Отделение экспериментальной гематологии, Кафедра клинической и экспериментальной медицины, Университет Линчёпинга, Линчёпинг, Швеция
4 Подразделение Инфекционная биология и медицинская микробиология, Департамент наук о жизни, Школа наук о жизни, Центральный университет Тамил Наду, Тируварур, Индия
5 Департамент исследований ревматологии и воспаления, Гетеборгский университет, Гетеборг, Швеция
Отредактировал: Карл ГРАММ. Фен, Сиднейский университет, Австралия
Фен, Сиднейский университет, Австралия
Рецензент: Энтони Лоуренс Каннингем, Институт медицинских исследований Вестмида, Австралия; Брайан МакШарри, Университетский колледж Корка, Ирландия
Эта статья была отправлена в Viral Immunology, раздел журнала Frontiers in Immunology
‡ Текущий адрес: Элиза Криски, Департамент здоровья населения и патобиологии, Колледж ветеринарной медицины, штат Северная Каролина. University, Raleigh, NC, United States
† Эти авторы внесли равный вклад в эту работу
Поступила в редакцию 13 августа 2019 г .; Принята в печать 25 ноября 2019 г.
Авторские права © 2019 Crisci, Svanberg, Ellegård, Khalid, Hellblom, Okuyama, Bhattacharya, Nyström, Shankar, Eriksson and Larsson. Это статья в открытом доступе, распространяемая в соответствии с условиями лицензии Creative Commons Attribution License (CC BY). Использование, распространение или воспроизведение на других форумах разрешено при условии указания автора (авторов) и правообладателя (ов) и ссылки на оригинальную публикацию в этом журнале в соответствии с принятой академической практикой. Запрещается использование, распространение или воспроизведение без соблюдения этих условий.
Запрещается использование, распространение или воспроизведение без соблюдения этих условий.
Abstract
Генитальный герпес — распространенная инфекция, передаваемая половым путем, вызываемая вирусом простого герпеса 2 типа (ВПГ-2). Генитальный герпес значительно усиливает приобретение и передачу ВИЧ-1, создавая микросреду, которая поддерживает ВИЧ-инфекцию в организме хозяина. Дендритные клетки (ДК) представляют собой один из первых врожденных типов клеток, которые сталкиваются с ВИЧ-1 и ВПГ-2 в слизистой оболочке половых органов. Было показано, что инфекция ВПГ-2 модулирует ДК, делая их более восприимчивыми к ВИЧ-инфекции.Здесь мы исследовали потенциальные механизмы, лежащие в основе опосредованного HSV-2 увеличения инфицирования ВИЧ-1. Мы продемонстрировали, что присутствие ВПГ-2 усиливало продуктивную инфекцию ДК ВИЧ-1 и усиливало воспалительные и противовирусные реакции. Инфекция ВИЧ-1, усиленная ВПГ-2, требовала интактной ДНК ВПГ-2, но не активной репликации ДНК ВПГ-2. Кроме того, усиленная ВИЧ-инфекция ДК затрагивает путь cGAS-STING. Интересно, что мы не смогли увидеть никакого участия TLR2 или TLR3 или подавления инфекции за счет продукции IFN-β.Кондиционирование HSV-2 в двойно экспонированных DC снижает экспрессию белков IFI16, cGAS, STING и TBK1, которые связаны с передачей сигналов через путь STING. Двойное воздействие ВПГ-2 и ВИЧ-1 привело к снижению уровней некоторых факторов рестрикции ВИЧ-1, особенно SAMHD1, TREX1 и APOBEC3G. Активация пути STING в ДК под воздействием как ВПГ-2, так и ВИЧ-1, скорее всего, привела к протеолитической деградации факторов рестрикции ВИЧ-1 SAMHD1, TREX1 и APOBEC3G, что должно ослабить их нормальное ограничение ВИЧ-инфекции в ДК. .Это привело к их нормальному ограничению ВИЧ-инфекции в DC. Мы показали, что перепрограммирование HSV-2 клеточных сигнальных путей и уровней экспрессии белка в DC обеспечивает условия, в которых ВИЧ-1 может вызвать более продуктивную инфекцию в DC. В заключение, репрограммирование HSV-2 открывает DC для заражения ВИЧ-1 и создает микросреду, благоприятствующую передаче ВИЧ-1.
Кроме того, усиленная ВИЧ-инфекция ДК затрагивает путь cGAS-STING. Интересно, что мы не смогли увидеть никакого участия TLR2 или TLR3 или подавления инфекции за счет продукции IFN-β.Кондиционирование HSV-2 в двойно экспонированных DC снижает экспрессию белков IFI16, cGAS, STING и TBK1, которые связаны с передачей сигналов через путь STING. Двойное воздействие ВПГ-2 и ВИЧ-1 привело к снижению уровней некоторых факторов рестрикции ВИЧ-1, особенно SAMHD1, TREX1 и APOBEC3G. Активация пути STING в ДК под воздействием как ВПГ-2, так и ВИЧ-1, скорее всего, привела к протеолитической деградации факторов рестрикции ВИЧ-1 SAMHD1, TREX1 и APOBEC3G, что должно ослабить их нормальное ограничение ВИЧ-инфекции в ДК. .Это привело к их нормальному ограничению ВИЧ-инфекции в DC. Мы показали, что перепрограммирование HSV-2 клеточных сигнальных путей и уровней экспрессии белка в DC обеспечивает условия, в которых ВИЧ-1 может вызвать более продуктивную инфекцию в DC. В заключение, репрограммирование HSV-2 открывает DC для заражения ВИЧ-1 и создает микросреду, благоприятствующую передаче ВИЧ-1.
Ключевые слова: Коинфекция ВПГ-2, ВИЧ-1, ВПГ-2 и ВИЧ-1, дендритные клетки, иммунные ответы, сенсоры ДНК
Введение
Инфекция, вызванная вирусом иммунодефицита человека (ВИЧ), является заболеванием, передаваемым половым путем, которое, как сообщается поражает почти 37 миллионов человек во всем мире (1).Риск заражения ВИЧ-1 во время полового акта в три раза выше для лиц, инфицированных вирусом простого генитального герпеса типа 2 (ВПГ-2) (2, 3), и наибольший риск наблюдается у лиц, которые столкнулись с ВПГ-2 недавно (4). Многие люди с серопозитивной реакцией на ВПГ-2 не знают о своем инфекционном статусе, а выделение вируса и эпителиальные микроязвы, как известно, часто возникают у бессимптомных лиц. Более того, данные указывают на то, что противовирусное лечение не может полностью подавить инфекцию HSV-2 (5–7).Присутствие как ВИЧ-1, так и ВПГ-2 создает среду, в которой иммунные клетки, такие как дендритные клетки (ДК) и клетки Лангерганса (ЖК), могут подвергаться воздействию вирусов или вирусных факторов и обусловливаться ими. Интересно, что присутствие как HSV-2, так и HIV-1, по-видимому, в несколько раз увеличивает уровни ДНК ВИЧ в DC (8), LC (9) и макрофагах (10).
Интересно, что присутствие как HSV-2, так и HIV-1, по-видимому, в несколько раз увеличивает уровни ДНК ВИЧ в DC (8), LC (9) и макрофагах (10).
Усиление ВИЧ-инфекции за счет присутствия HSV-2, по-видимому, имеет несколько объяснений, включая микросреду, обусловленную инфильтрацией и персистенцией HSV-2-специфических CD4 + T-клеток в слизистой оболочке (11).CD4 + Т-клетки из-за их экспрессии α4β7 + особенно восприимчивы к ВИЧ-инфекции (12–14). Кроме того, повышенная инфильтрация миелоидных ДК и плазмоцитоидных ДК (12), вероятно, способствует усилению инфекции слизистой оболочки за счет привлечения дополнительных клеток-мишеней ВИЧ. Причина усиления инфекции, помимо большего количества присутствующих иммунных клеток, может быть связана с активацией ДК, расположенных в слизистой оболочке (15). Активированные DC продуцируют высокие уровни TNF, которые обусловливают микросреду, а также соседние DC и Т-клетки, делая их более восприимчивыми к инфекции ВИЧ-1 за счет увеличения их экспрессии CCR5 (8). Кроме того, HSV-2 может также предварительно кондиционировать DC и Т-клетки, обеспечивая патоген-ассоциированные молекулярные структуры (PAMP), что приводит к усилению ВИЧ-инфекции.
Кроме того, HSV-2 может также предварительно кондиционировать DC и Т-клетки, обеспечивая патоген-ассоциированные молекулярные структуры (PAMP), что приводит к усилению ВИЧ-инфекции.
Микроокружение в очаге инфекции обогащено набором растворимых факторов, которые могут влиять на инфицирование ВИЧ-1 и ВПГ-2, включая компоненты системы комплемента (16–18). Вирионы ВИЧ-1 защищены от нормального лизиса, опосредованного комплементом, благодаря включению молекул, ингибирующих комплемент, таких как CD55 и CD59, в их липидную мембрану (19), что приводит к опсонизации частиц ВИЧ-1 инактивированными фрагментами комплемента, такими как iC3b и C3d (20–22).Кроме того, вирионы, передаваемые во время половой передачи, должны быть комплементарными и / или комплементарными частицами, опсонизированными антителами, видимыми при контакте с семенной жидкостью и / или секрециями шейки матки (19). Считается, что оба вируса используют это звено врожденного иммунного аппарата для усиления тяжести инфекции и избегания комплемент-опосредованного лизиса вирионов (18, 20, 21, 23).
Активация врожденного иммунитета является важным компонентом механизмов противовирусной защиты. Пока что о врожденном клеточном программировании, генерируемом HSV-2 в DC, известно значительно меньше, что может способствовать развитию ВИЧ-инфекции.И HSV-2, и ВИЧ распознаются множеством рецепторов распознавания образов (PRR) в организме хозяина. Ранее были показаны Toll-подобный рецептор (TLR) 2, TLR3, TLR9 и некоторые внутрицитоплазматические сенсоры, такие как отсутствующие в меланоме 2 (AIM2), интерферон-гамма-индуцибельный белок 16 (IFI16) и белок 5, связанный с дифференцировкой меланомы (MDA5). для распознавания HSV в различных типах клеток мыши и / или человека (24, 25). Раннее распознавание HSV-1 в первичных клетках человека, например, макрофагах, включает два внутриклеточных пути восприятия нуклеотидов для индукции IFN типа I и воспалительных факторов.MDA5 / MAVS играет первостепенную роль в индукции IFN типа I, тогда как путь TLR2, по-видимому, играет менее важную роль в ранних врожденных ответах (24). Недавние открытия установили участие врожденных сенсоров IFI16 и cGMP-AMP-синтазы (cGAS) в защите хозяина от ДНК-вирусов, таких как HSV (26). Восприятие IFI16 ДНК ВПГ активирует инфламмасомный аппарат и ответы IFN-β в некоторых типах клеток (26). В фибробластах, инфицированных HSV-1, ДНК HSV-1 воспринимается как IFI16, так и cGAS (27).IFI16 и cGAS стимулируют путь STING, что приводит к активации TBK1 и IRF3, что приводит к продукции IFN-β (28). ВИЧ-1 распознается TLR, RIG-1-подобными рецепторами, NOD2, IFI16 и cGAS (29–31). Мы обнаружили, что передача сигналов в ВИЧ-инфицированных ДК, происходящих из моноцитов человека, включает путь TLR8 с последующей активацией сигнальных каскадов p38 MAPK, ERK, NF-κB, IRF1 и IRF7, тогда как опсонизированные комплементом вирионы обладают способностью подавлять эти и активируют сигнальные пути Lyn, PI3K, IRF3 (20).
Недавние открытия установили участие врожденных сенсоров IFI16 и cGMP-AMP-синтазы (cGAS) в защите хозяина от ДНК-вирусов, таких как HSV (26). Восприятие IFI16 ДНК ВПГ активирует инфламмасомный аппарат и ответы IFN-β в некоторых типах клеток (26). В фибробластах, инфицированных HSV-1, ДНК HSV-1 воспринимается как IFI16, так и cGAS (27).IFI16 и cGAS стимулируют путь STING, что приводит к активации TBK1 и IRF3, что приводит к продукции IFN-β (28). ВИЧ-1 распознается TLR, RIG-1-подобными рецепторами, NOD2, IFI16 и cGAS (29–31). Мы обнаружили, что передача сигналов в ВИЧ-инфицированных ДК, происходящих из моноцитов человека, включает путь TLR8 с последующей активацией сигнальных каскадов p38 MAPK, ERK, NF-κB, IRF1 и IRF7, тогда как опсонизированные комплементом вирионы обладают способностью подавлять эти и активируют сигнальные пути Lyn, PI3K, IRF3 (20).
В свете повышенной распространенности ВИЧ-инфекции и выделения вируса на слизистую оболочку половых органов у серопозитивных людей по ВПГ-2 (2, 3, 5-7), а также того факта, что ДК становятся мишенью для ВИЧ-1 во время передачи половым путем, мы стремились расшифровать индуцированное ВПГ-2 клеточное программирование, лежащее в основе усиленной инфекции ДК ВИЧ-1. Предварительное кондиционирование человеческих DC HSV-2 перед воздействием ВИЧ-1 увеличивало воспалительные и противовирусные реакции, а также уровень ВИЧ-инфекции в DC. Мы обнаружили, что множественные сигнальные пути активируются на уровне транскрипции как в свободных, так и в опсонизированных комплементом DCs, подвергнутых воздействию HSV-2 / HIV-1.Усиление инфицирования ВИЧ-1 зависело от активации пути STING и требовало распознавания структурно интактной ДНК HSV-2. Кондиционирование HSV-2, как свободным, так и опсонизированным комплементом, в двойно экспонированных DC снижает уровни белков IFI16, cGAS, TBK1 и IRF3, которые все участвуют в пути STING. Кроме того, активация пути STING в DC, инфицированных HSV-2 / HIV-1, привела к снижению уровней некоторых факторов рестрикции ВИЧ, например SAMHD1, TREX1 и APOBEC3G, что уменьшило их клеточное ограничение инфекции ВИЧ-1 в DC.Наша работа продемонстрировала, что репрограммирование, опосредованное ВПГ-2, увеличивает способность ВИЧ инфицировать ДК и создает благоприятную микросреду для инфицирования и передачи ВИЧ.
Предварительное кондиционирование человеческих DC HSV-2 перед воздействием ВИЧ-1 увеличивало воспалительные и противовирусные реакции, а также уровень ВИЧ-инфекции в DC. Мы обнаружили, что множественные сигнальные пути активируются на уровне транскрипции как в свободных, так и в опсонизированных комплементом DCs, подвергнутых воздействию HSV-2 / HIV-1.Усиление инфицирования ВИЧ-1 зависело от активации пути STING и требовало распознавания структурно интактной ДНК HSV-2. Кондиционирование HSV-2, как свободным, так и опсонизированным комплементом, в двойно экспонированных DC снижает уровни белков IFI16, cGAS, TBK1 и IRF3, которые все участвуют в пути STING. Кроме того, активация пути STING в DC, инфицированных HSV-2 / HIV-1, привела к снижению уровней некоторых факторов рестрикции ВИЧ, например SAMHD1, TREX1 и APOBEC3G, что уменьшило их клеточное ограничение инфекции ВИЧ-1 в DC.Наша работа продемонстрировала, что репрограммирование, опосредованное ВПГ-2, увеличивает способность ВИЧ инфицировать ДК и создает благоприятную микросреду для инфицирования и передачи ВИЧ.
Материалы и методы
Реагенты
В питательную среду DC RPMI1640 (GIBCO, Швеция) добавляли 2 мМ глутамина, 20 мкг / мл гентамицина (GIBCO), 10 мМ HEPES (GIBCO) и 1% человеческой плазмы. Рекомбинантный человеческий GM-CSF (Genzyme) (100 Ед / мл) и 300 Ед / мл рекомбинантный человеческий ИЛ-4 (R&D Systems, Миннеаполис, Миннесота, США) использовали для размножения DC in vitro .
Полученные из моноцитов DC и культура клеток THP1
Была собрана цельная кровь здоровых добровольцев или лейкоцитарная пленка из банка крови в университетской больнице Линчёпинга (этические разрешения M173-07 и M75-08 / 2008). Мононуклеарные клетки периферической крови (PBMC) разделяли центрифугированием в градиенте плотности с использованием Ficoll-Hypaque (Amersham Pharmacia Biotech, Piscataway, NJ, USA) и инкубировали на чашках для культивирования клеток (BD, Европа) в течение 1 часа при 37 ° C, чтобы обеспечить прилипание Предшественники DC и возможность отбрасывать неприлипающие клетки. Предшественники были дифференцированы в незрелые ДК, происходящие из моноцитов (далее называемые незрелыми ДК), путем добавления 100 ед. / Мл GM-CSF и 300 ед. / Мл IL-4 на 0, 2 и 4 день культивирования. После этого DC оценивали на экспрессию маркеров CD14 и CD83 в качестве контроля качества перед использованием в экспериментах. В некоторых экспериментах использовали клетки THP1 дикого типа или THP1-Dual ™ KO-STING (Invivogen, Франция). Клетки THP1 культивировали в соответствии с инструкциями производителя, активировали, используя форбол 12-миризат 13-ацетат (PMA, 10 мкг / мл), и инкубировали за 2 дня до инфицирования клеток и обрабатывали таким же образом, как описано ниже для DC.
Предшественники были дифференцированы в незрелые ДК, происходящие из моноцитов (далее называемые незрелыми ДК), путем добавления 100 ед. / Мл GM-CSF и 300 ед. / Мл IL-4 на 0, 2 и 4 день культивирования. После этого DC оценивали на экспрессию маркеров CD14 и CD83 в качестве контроля качества перед использованием в экспериментах. В некоторых экспериментах использовали клетки THP1 дикого типа или THP1-Dual ™ KO-STING (Invivogen, Франция). Клетки THP1 культивировали в соответствии с инструкциями производителя, активировали, используя форбол 12-миризат 13-ацетат (PMA, 10 мкг / мл), и инкубировали за 2 дня до инфицирования клеток и обрабатывали таким же образом, как описано ниже для DC.
HSV-2, запас вируса готовили в клетках почки африканской зеленой мартышки (GMK), культивируемых в DMEM с добавлением 10% инактивированной нагреванием (HI) FCS, как описано ранее (32). Штамм 333 HSV-2 использовали либо как инфекционный, либо как γ-облученный (30 мин) инактивированный вирус. ВИЧ-1BaL / SUPT1-CCR5 CL.30 (партии 4235, 4238, 4313 и 4366) получали с использованием хронически инфицированных культур линии клеток ACVP / BCP (№ 204), первоначально полученных путем инфицирования SUPT1-CCR5 CL.30 клеток (щедро подаренных доктором Дж. Хокси из Пенсильванского университета) с инфекционным запасом ВИЧ-1BaL (Программа исследований и эталонных реагентов NIH AIDS, Каталожный № 416, Лот № 59155). Вирус очищали и концентрировали, как описано ранее (33), и аликвоты замораживали. Все вирусные препараты проверяли на инфекционность.
ВИЧ-1BaL / SUPT1-CCR5 CL.30 (партии 4235, 4238, 4313 и 4366) получали с использованием хронически инфицированных культур линии клеток ACVP / BCP (№ 204), первоначально полученных путем инфицирования SUPT1-CCR5 CL.30 клеток (щедро подаренных доктором Дж. Хокси из Пенсильванского университета) с инфекционным запасом ВИЧ-1BaL (Программа исследований и эталонных реагентов NIH AIDS, Каталожный № 416, Лот № 59155). Вирус очищали и концентрировали, как описано ранее (33), и аликвоты замораживали. Все вирусные препараты проверяли на инфекционность.
Генерация репортера GFP CCR5-Tropic Virus
Провирусная конструкция NLENG1-IRES была использована для создания NLENG1-IRES-70 путем замены ENV на YU-2 ENV, как описано в другом месте (34, 35).Провирусная конструкция была щедро пожертвована доктором Дэвидом Н. Леви (Нью-Йоркский университет, Нью-Йорк, штат Нью-Йорк, США). Клетки HEK-293T культивировали в среде DMEM, содержащей 10% HI FCS, и при ~ 70% конфлюентности клетки трансфицировали провирусной конструкцией NLENG1-IRES-70 с использованием метода CaPO 4 . После 8 ч трансфекции среду заменяли DMEM с добавлением 1% HI FCS. GFP-HIV собирали на следующий день, собирая супернатант, и остатки клеток удаляли осаждением при 2500 об / мин в течение 5 минут.Исходные вирусы были разделены на аликвоты и заморожены при -80 ° C.
После 8 ч трансфекции среду заменяли DMEM с добавлением 1% HI FCS. GFP-HIV собирали на следующий день, собирая супернатант, и остатки клеток удаляли осаждением при 2500 об / мин в течение 5 минут.Исходные вирусы были разделены на аликвоты и заморожены при -80 ° C.
Опсонизация HSV-2 и ВИЧ
Комплементарная опсонизация HSV-2, HIV и GFP-HIV была проведена путем инкубации вирионов с равным объемом человеческой сыворотки (HS) (этические разрешения M173-07 и M75 -08/2008). Для опсонизации использовались разные HS; Серонегативный HSV-1 и HSV-2 был использован для опсонизации вируса HSV-2 (обозначаемого как CHSV), а серонегативный HS-ВИЧ и HSV-2 был использован для опсонизации вирусов ВИЧ-1 или GFP-HIV-1 (обозначаемых как CHIV и CHIV-GFP).HS был протестирован на антитела к HSV с использованием наборов HerpeSelect® 1 ELISA IgG и HerpeSelect® 2 ELISA IgG (Focus Diagnostics, Cypress, CA, USA). Свободные вирусы HSV-2, HIV-1 и GFP-HIV-1 (называемые HSV, HIV и GFP-HIV) разводили до той же концентрации опсонизированного вируса. Отрицательный контроль, т.е. имитация, обрабатывали культуральной средой. Все вирусные группы инкубировали в течение 1 ч при 37 ° C и использовали в экспериментах по коинфекции.
Отрицательный контроль, т.е. имитация, обрабатывали культуральной средой. Все вирусные группы инкубировали в течение 1 ч при 37 ° C и использовали в экспериментах по коинфекции.
HSV-2 и ВИЧ-инфекция DC
Незрелые DC (10 6 клеток / мл) были инфицированы ложным, (30 нг / мкл) ВИЧ или CHIV как единичная инфекция.ВИЧ / HSV или CHIV / CHSV использовались в двух схемах: прямая и отсроченная коинфекция. В отсроченной группе 1 MOI HSV-2 и CHSV добавляли к DC в течение 2 часов, а затем клетки промывали перед добавлением ВИЧ и CHIV в течение 4, 6 или 22 часов. В установке прямого совместного инфицирования оба типа вируса добавляли к DC одновременно в течение 3 часов, промывали и затем инкубировали еще 21 час во всех установках. DC культивировали в среде 1% плазмы человека. ДК собирали, промывали и лизировали лизисным буфером Bioline RLY (Bioline, UK) для экстракции РНК или фиксировали 4% параформальдегидом (PFA) в течение 10 минут при 4 ° C для иммунного фенотипирования с помощью проточной цитометрии.
Чтобы изучить уровень продуктивной ВИЧ-инфекции, были выполнены две схемы коинфекции, и супернатанты были собраны на 1, 4, 7 и 10 день. После сбора супернатанта клетки повторно суспендировали. в 10% FBS перед добавлением IL-4 и GM-CSF. Уровень p24 ВИЧ определяли с помощью собственного анализа ELISA на p24 (36). Кроме того, продуктивная ВИЧ-инфекция оценивалась с использованием репортерного CCR5-тропного вируса GFP с использованием тех же экспериментальных установок, которые описаны ранее для других вирусов.DC инфицировали GFP-HIV в течение 3 и 5 дней перед тем, как процент положительных клеток GFP анализировали с помощью проточной цитометрии.
Экстракция общей РНК, обратная транскрипция и кПЦР
Общая РНК из ДК, подвергшихся воздействию различных условий, экстрагировалась с использованием коммерческого набора Isolate II RNA Mini или Micro (Bioline, UK), а общая кДНК была получена с помощью обратной транскриптазы SuperScript III Набор для синтеза первой цепи кДНК (Invitrogen, Carlsbad, CA, USA). Количественную оценку транскриптов генов проводили с использованием набора SensiFAST SYBR® Hi-ROX (Bioline, UK) и системы CFX96 Touch Real-Time (BIO-RAD Inc.). Праймеры, нацеленные на β-актин и GAPDH, использовали в качестве генов домашнего хозяйства для справки, как описано другими (37). Праймеры были закуплены у CyberGene AB (Стокгольм, Швеция). Чтобы компенсировать различия между пластинами, значения были нормализованы, как описано ранее (38).
Количественную оценку транскриптов генов проводили с использованием набора SensiFAST SYBR® Hi-ROX (Bioline, UK) и системы CFX96 Touch Real-Time (BIO-RAD Inc.). Праймеры, нацеленные на β-актин и GAPDH, использовали в качестве генов домашнего хозяйства для справки, как описано другими (37). Праймеры были закуплены у CyberGene AB (Стокгольм, Швеция). Чтобы компенсировать различия между пластинами, значения были нормализованы, как описано ранее (38).
ELISA и Cytometric Bead Array
Уровни белков TNF (Mabtech, Швеция) и IFN-β (набор VeriKine; PBL Assay Science, США) оценивали в супернатантах культур DC примерно через 20 часов инфицирования с помощью ELISA в соответствии с протоколы производителей.Наборы цитометрических шариков (BD Biosciences, Стокгольм, Швеция) использовали для измерения уровней цитокинов и хемокинов в клеточных супернатантах и для измерения уровней фосфорилированного белка в лизатах DC (BD Biosciences, Стокгольм, Швеция) в соответствии с протоколами производителя.
Проточная цитометрия
Качество незрелых ДК оценивали путем окрашивания антителами, конъюгированными с PE человека против CD83 и CD14 (BD, Европа). DC использовали, если чистота была> 95% и их экспрессия CD14 и CD83 была <10%.Экспрессию антигенов HSV-2 в DC, подвергнутых воздействию HSV-2, осуществляли с помощью поликлональных антител против HSV-2 (B0116, Dako, Дания). Влияние инфекции на различные белки оценивали путем окрашивания конъюгированным с PE-Cy7 антителом против APOBEC3G человека (Abcore, США), конъюгированным с Alexa Fluor 647 антителом против SAMHD1 человека (Bioss Inc., США), конъюгированным с Alexa Fluor 647 анти- человеческий TREX1, антитела против IRF3 человека, конъюгированные с PE антитела против NAK / TBK1 человека, антитела против человеческого IFI16 (Abcam, Великобритания), против CF150 человека (cGAS) (Thermofisher Scientific) и конъюгированные с PE антитела против STING человека ( BD Europe).Неконъюгированные антитела окрашивали конъюгированными вторичными антикроличьими антителами (DAKO). Окрашенные образцы DC оценивали с помощью проточной цитометрии (FACS Canto II, BD) и анализировали с помощью FlowJo (Treestar, Ashland, OR, США).
Лиганды и ингибиторы
В экспериментах использовались следующие лиганды и ингибиторы TLR: лиганд TLR2 PAM (5 мкг / мл), лиганд TLR3 Poly I: C (40 мкг / мл), 2′3′-cGAMP, ( 10 мкг / мл), Poly dA: dT (0,5 мкг / мл), голая ДНК HSV-60 (10 мкг / мл), олигонуклеотид A151 ODN TTAGGG (1-3 мкМ) (Invivogen, Франция), pp65 человека (UL83) рекомбинантный белок (10–15 мкг / мл) (Miltenyi Biotec, Швеция), рекомбинантный белок E7 вируса папилломы человека 18 типа (10–20 мкг / мл) (SMS-gruppen, Дания) и ацикловир (20–50 мкМ) (SIGMA ).Ацикловир добавляли за 1 час до добавления вируса, инкубировали с вирусом в течение 2 часов, смывали, пополняли и после этого оставляли в культуре на протяжении всей инкубации. Внутриклеточная доставка cGAMP, Poly dA: dT, голой ДНК HSV-60, олигонуклеотида A151, белка UL83 и белка E7 была достигнута с использованием коммерческой системы DOTAP (SIGMA). Вкратце, различные лиганды и ингибиторы смешивали с забуференным HEPES физиологическим раствором и DOTAP в течение 30 минут при комнатной температуре, инкубировали в течение 2 часов при 37 ° C с DC и промывали перед инфицированием.Только ДОТАП служил контролем за действием системы доставки.
RNAseq
РНК (5 нг) подвергали амплификации полного транскриптома с использованием набора NuGEN Ovation RNA-Seq V2 (Сан-Карлос, Калифорния, США) в соответствии с инструкциями производителя. Вкратце, кДНК амплифицировали из общей РНК с использованием изотермической амплификации с одним праймером (SPIA). Затем амплифицированные образцы кДНК очищали с использованием набора MinElute Reaction Cleanup Kit (Qiagen; Валенсия, Калифорния, США). Образцы кДНК были фрагментированы на более мелкие части, с тупыми концами, лигированы с индексированными (штрих-кодированными) адаптерами и амплифицированы с использованием набора Ultralow System V2 в соответствии с протоколом производителя.Окончательное распределение размеров библиотек определяли с использованием Agilent Bioanalyzer 2100. Пять библиотек от пяти разных доноров секвенировали на платформе Illumina NextSeq500 (Сан-Диего, Калифорния, США). Файлы FASTQ были загружены в UPPMAX и качество проверено с помощью fastQC (39). Trimmomatic (40) использовался для удаления адаптеров и низкокачественных оснований, и считанные данные затем были сопоставлены с эталонным геномом человека hg19 с помощью STAR (41). Количество для каждого гена было рассчитано с помощью featureCounts (42). Данные были нормализованы и дифференциально экспрессируемые гены определены с использованием R / DeSeq2 (43).Анализ путей проводился с помощью анализа пути изобретательности (Qiagen), анализа R, анализа обогащения онтологией генов (GO) (Geneontology.org) и пользовательских списков генов.
Статистический анализ
GraphPad Prism 5 (GraphPad Software, La Jolla, CA, USA) использовали для анализа всех данных, кроме RNAseq. Для проверки статистической значимости использовали повторные измерения ANOVA с последующим пост-тестом Бонферрони или двусторонний парный тест t . Результаты считались статистически значимыми, если p <0.05. Все эксперименты проводились минимум четыре раза с использованием клеток, полученных от разных доноров крови. Когда экспериментальные значения были нормализованы, среднее значение свободного вируса или ложного вируса было установлено на 1. Результаты qPCR были нормализованы для различий между планшетами, как описано ранее (38). Вкратце, каждое значение было вычтено из среднего значения всех значений, а затем полученные значения были разделены на среднее значение ложного или свободного вируса в зависимости от различных экспериментов.
Результаты
Кондиционирование ВПГ-2 человеческих ДК сделало их более восприимчивыми к ВИЧ-инфекции
Теперь очевидно, что инфекция половых путей ВПГ-2 предрасполагает к ВИЧ-инфекции и увеличению выделения вируса на слизистую оболочку (2, 3 , 5–7).Тем не менее механизмы, лежащие в основе повышенной инфекции, остаются неоднозначными. Здесь мы оценили роль ДК в усилении инфекции, оценивая эффекты до воздействия ВПГ-2 на ВИЧ-инфекцию человеческих ДК, происходящих из моноцитов, и роль опсонизации вирусов комплементом. Уровни транскриптов мРНК gag ВИЧ были значительно увеличены в ДК, подвергнутых двойному воздействию ВПГ / ВИЧ, по сравнению с уровнями, индуцированными только ВИЧ, как через 6, так и через 24 часа (). Связанное с HSV-2 усиление ВИЧ-инфекции согласуется с выводами Marsden et al.(8). Более того, DC, подвергнутые воздействию опсонизированного комплементом HSV-2, давали более высокий тирозин-кинез (TK) как в двойном, так и в однократном воздействии DC по сравнению со свободным вирусом, как показано ранее, но не было изменений в инфекции HSV-2 при измерении экспрессии TK. в DC, подвергнутых двойному воздействию ВИЧ / HSV, по сравнению с DC, подвергнутым однократному воздействию HSV-2 (дополнительный рисунок 1). Продуктивная ВИЧ-инфекция также была значительно повышена в клетках, подвергнутых двойному воздействию ВПГ / ВИЧ, что измерено процентным содержанием GFP-ВИЧ-положительных ДК с помощью проточной цитометрии и конфокальной микроскопии на 3-й и 5-й день после контакта (), а также уровнями p24 ВИЧ в культуре. супернатанты день 4 и день 1–9 ().Опсонизация вирионов существенно не изменила уровень ВИЧ-инфекции в клетках, инфицированных ВПГ / ВИЧ (). Следует отметить, что присутствие HSV-2 усиливало ВИЧ-инфекцию, как когда DC были предварительно кондиционированы HSV-2 перед ВИЧ-инфекцией, так и когда HSV-2 и ВИЧ были введены в DC одновременно. В нормальных условиях только небольшая часть DC будет продуктивно инфицирована ВИЧ (), то же самое относится и к HSV-2 (18). Тем не менее, все ДК в культурах подвергаются воздействию вирусов и подвергаются их влиянию либо косвенно, либо напрямую, даже в отсутствие продуктивной инфекции (18, 36).Количество ДК, поддерживающих продуктивную инфекцию как ВИЧ, так и ВПГ-2, невелико и было показано Marsden et al. чтобы представить только 1% ячеек. В нашей системе мы определили, что уровень DC с продуктивной ВИЧ-инфекцией и положительным по антигену HSV-2 должен составлять 5% для DC, подвергнутого свободному воздействию HIV / HSV, и ~ 12% для DC, подвергнутого воздействию опсонизированного комплементом CHIV / CHSV (), что может отражают, что не все антиген-положительные клетки HSV-2 продуктивно инфицированы, скорее, что некоторые клетки имеют абортивную инфекцию HSV (44–46).Данные для прямой и отсроченной коинфекции дали очень похожие данные, поэтому эксперименты, представленные и проведенные далее, основаны на установке отсроченной коинфекции. Взятые вместе, наши данные ясно демонстрируют, что воздействие HSV-2 увеличивает способность ВИЧ продуктивно инфицировать DC, что побудило нас выяснить основной механизм, ответственный за усиление ВИЧ-инфекции.
ВПГ-2 кондиционирование дендритных клеток человека сделало их более восприимчивыми к ВИЧ-продуктивной ВИЧ-инфекции. Дендритные клетки подвергали воздействию свободного HSV-2 (HSV) или опсонизированного комплементом вируса (CHSV) в течение 2 часов, затем промывали и инфицировали свободным ВИЧ, либо ВИЧ-1 BaL, либо GFP-меченный CCR5 тропический ВИЧ-1 (GFP-HIV) или комплемент опсонизированный ВИЧ (CHIV) в течение 6 или 22 часов.Положительные ВПГ-2 DC измеряли с использованием поликлональных антител против ВПГ-2 с последующим измерением конъюгированных с флуорохромом вторичных антител (А). Уровни экспрессии мРНК HIV gag определяли с помощью количественной ПЦР через 6 и 24 часа. Данные нормализуются для бесплатного вируса, который устанавливается как один. (B) GFP-ВИЧ-положительные клетки оценивали с помощью проточной цитометрии через 3 и 5 дней после инфицирования ВИЧ или с помощью конфокальной микроскопии (40 ×) через 5 дней после заражения. GFP-HIV продуктивно инфицированные клетки (зеленые), окрашенные ядерным окрашиванием DAPI (синий).Размерная полоса = 20 мкм. (C) Высвобождение p24 ВИЧ-1 продуктивно инфицированными DC определяли с помощью ELISA с 1 по 9 день после инфицирования ВИЧ. (D) Процент ВИЧ-GFP-положительных и антиген-положительных ВПГ-2 DC, измеренный с помощью проточной цитометрии через 24 часа после инфицирования. * p <0,05; ** р <0,005; *** р <0,0005. N = 5–8.
Различное клеточное программирование было индуцировано в ДК различными условиями инфицирования ВИЧ и ВПГ-2
Для дальнейшего понимания клеточного программирования во время инфекции ВПГ / ВИЧ мы выполнили секвенирование РНК, которое обеспечило полный профиль транскриптома в ДК, индуцированных одним контакт с ВИЧ vs.двойное воздействие ВПГ-2 и ВИЧ. Анализ транскриптома продемонстрировал активацию и участие различных наборов путей в одиночных и двойных инфицированных DC соответственно. ДК, подвергнутые двойному воздействию вируса, активировали или подавляли значительно больше генов, чем ДК, подверженные только ВИЧ и CHIV (). Это продемонстрировало, что воздействие как ВПГ, так и ВИЧ было более сильным активатором ДК, чем только ВИЧ, и что опсонизация комплемента вирионов подавляла активацию ДК. Затем наш анализ обогащения генов четко установил, что каждое из различных вирусных состояний приводит к уникальным паттернам экспрессии генов в различных сигнальных путях (; дополнительный рисунок 2).В условиях CHIV и CHIV / CHSV были менее обогащенные пути по сравнению с группами свободных вирусов (). Гены в путях передачи сигналов интерлейкина и JAK / STAT были обогащены как ДК, подвергнутым двойному воздействию, так и ВИЧ, CHIV и ВИЧ / ВПГ, но только ВИЧ / ВПГ имели значительное обогащение (дополнительный рисунок 2). Недавно было показано, что путь убиквитин-протеасома играет важную роль в вирусном патогенезе (47). Гены, участвующие в этом пути, были значительно обогащены свободными контактами ВИЧ и ВИЧ / ВПГ, но только ограниченное количество генов было обогащено в группах, подвергшихся воздействию комплемента (дополнительный рисунок 2).Обогащенный анализ генов со сниженным уровнем экспрессии после ВИЧ-инфекции показал, что гены в пути биосинтеза холестерина были значительно обогащены ДК, подвергнутыми воздействию CHIV, а гены, участвующие в метаболизме пиримидина, были обогащены как ДК, подверженными воздействию ВИЧ / ВПГ, так и CHIV / CHSV (дополнительные Фигура 2). Более конкретный, углубленный анализ обогащения генов с акцентом на воспаление, противовирусные и иммунные ответы выявил паттерн значительно обогащенных путей, отчетливо различающихся для каждого типа инфекционного состояния, хотя между группами было некоторое совпадение (; дополнительный рисунок 2) .Например, активированные гены в каскаде JAK-STAT были значительно обогащены для всех групп, кроме CHIV (дополнительный рисунок 2).
Клеточное программирование дендритных клеток при одиночной ВИЧ-инфекции и двойной инфекции ВИЧ / ВПГ. Дендритные клетки подвергали воздействию HSV-2 (HSV) или опсонизированного комплементом вируса (CHSV) в течение 2 часов, затем инфицировали ВИЧ или опсонизированным комплементом ВИЧ (CHIV) в течение 24 часов. Было выполнено полное секвенирование транскриптома. (A) Анализ количества значительно активированных или подавляемых генов с оценкой общих, двух-, четырех- и восьмикратных изменений по сравнению с ложным (B) Анализ обогащения генов для генов, значительно активированных в три или более раз с помощью p ≥ 0.05 с акцентом на воспалительные и противовирусные пути / факторы. (C) Анализ обогащения генов значительно активизировал в три раза или выше с p ≥ 0,05. Анализ обогащения GO был проведен с набором данных о путях PANTHER. Термины со статистической значимостью в любом списке генов показаны в виде столбчатой диаграммы с накоплением. * p <0,05, ** p <0,005, *** p <0,0005. Ось Y = количество перечисленных генов, участвующих в указанном пути PANTHER.Ссылка = ожидаемый номер гена из ссылки (полные гены человека в базе данных). N = 5.
Воспалительные факторы были сильно повышены при двойном воздействии на ВПГ-2 и ВИЧ
Известно, чтоВИЧ и ВПГ-2 индуцируют воспалительные пути в иммунных клетках, причем ВПГ-2 является более сильным активатором (18 , 20), и наш анализ транскриптома ясно продемонстрировал активацию и участие нескольких воспалительных путей. HSV / HIV или CHSV / CHIV индуцировали различные паттерны экспрессии генов в DC для воспалительных генов по сравнению с одной инфекцией HIV или CHIV ().Более того, опсонизированные комплементом группы имели более низкие уровни транскрипции многих воспалительных факторов (). Один только ВИЧ, HSV / HIV и CHSV / CHIV активировали несколько воспалительных факторов, таких как IL-6, TNF, CXCL1 и CCL20, причем самая высокая экспрессия активировалась в группах с коинфекцией. Опсонизация комплемента снижает транскрипцию нескольких факторов, таких как CCL1, CCL5, CCL8 и IL-1β, видя, что они были ниже в опсонизированных группах по сравнению с условиями без вируса. CCL21, CCL28 и IL34 были активированы только в группах, подвергнутых двойному воздействию.Некоторые гены имели более высокие уровни экспрессии в группе CHIV и включали CXCR3 и IL-10. Затем мы проверили характер экспрессии некоторых воспалительных факторов с помощью кПЦР и трансляции с помощью ELISA или набора гранул цитокинов на основе потока. Сам по себе HSV был ведущим индуктором TNF и CCL3, тогда как ВИЧ-1 был более мощным в отношении индукции CXCL8 и IL-1β (дополнительная фигура 3). HSV / HIV и CHSV / CHIV приводили к более высоким уровням экспрессии мРНК TNF, CXCL8 и CCL3 по сравнению со свободным и опсонизированным комплементом ВИЧ ().Уровни мРНК IL-1β были одинаковыми в DC, подвергнутых воздействию ВИЧ и ВИЧ / HSV (), тогда как уровни белка были выше в DC, подвергнутых двойному воздействию (). Секреция TNF и хемокинов CCL3 и CCL5 следовала паттерну экспрессии мРНК и / или данным транскриптома (данные не показаны). Уровни белка других воспалительных факторов, таких как CXCL10, были ниже в DC, подвергшихся воздействию обоих вирусов, по сравнению с одним ВИЧ (). Эти несоответствия между уровнями белка и данными транскриптома могут быть объяснены посттранскрипционной регуляцией и / или протеолитической деградацией (48-50).Транскрипция воспалительных факторов активируется несколькими сигнальными путями, включая путь MAPK p38, и двойное воздействие значительно увеличивает уровень фосфорилированного p38 с тенденцией, что опсонизация вирионов снижает фосфорилирование p38 (2). Фосфорилирование JNK показало тот же паттерн активации, что и MAPK p38, но не было никаких существенных различий (данные не показаны), что могло быть связано с моментом времени, выбранным для измерения.
Воспалительные пути были сильно активированы в DC, подвергнутых воздействию HSV / HIV, на транскрипцию мРНК. (A) Дендритные клетки подвергали воздействию свободного HSV-2 или опсонизированного комплементом вируса (CHSV-2) в течение 2 часов, затем инфицировали ВИЧ или опсонизированным комплементом ВИЧ (CHIV) в течение 22 часов. Тепловая карта по воспалительным факторам, значительно повышенная или пониженная в одной или нескольких группах DC HIV, CHIV, HSV-2 / HIV или CHSV-2 / CHIV, по сравнению с ложно обработанными DC из экспрессии мРНК seq (B) РНК уровни IL-1β, TNF, CCL3, CXCL8 определяли с помощью ПЦР. Данные были нормализованы к ложному набору 1. (C) Уровни секретируемых цитокинов оценивали с помощью ELISA для TNF и набора гранул цитокинов для IL-1β, CCL5 и CXCL10. (D) Уровень фосфорилирования MAPK p38 оценивали в лизатах из DC, подвергнутых воздействию HSV-2 или CHSV-2 в течение 2 часов с последующим воздействием ВИЧ или CHIV в течение 4 часов с помощью набора фосфопротеиновых шариков. * p <0,05; ** р <0,005; *** р <0,0005. N = 5–8.
Воздействие ВИЧ / ВПГ вызывало более высокую экспрессию нескольких противовирусных факторов, включая IFN-β, в ДК по сравнению с воздействием только ВИЧ
Анализ транскриптома () четко продемонстрировал активацию и участие множества противовирусных путей в ДК, подвергшихся воздействию только ВИЧ, а также и ВПГ, и ВИЧ.Здесь мы оценили факторы, участвующие в активации и регуляции клеточной противовирусной защиты (). Паттерн для многих противовирусных факторов, индуцированных в DC, был аналогичен между ВИЧ и условиями двойных вирусных DC и в целом более низким уровнем антивирусных факторов, активированных в группах комплемента. Несколько генов TRIM, гены IFN типа I, гены SOCS и гены IRF были активированы в свободном ВИЧ и в двойно экспонированных DC, при этом самые высокие уровни транскрипции, в основном, обнаруживались в двойно экспонированных DC.Многие антивирусные гены были активированы в DC, подвергнутых воздействию HIV / HSV и CHIV / CHSV, тогда как было обнаружено, что несколько антивирусных генов активированы только в DC, подверженных воздействию ВИЧ, и включают IFITM3, SOCS5 и TBK1 для свободного ВИЧ и IFITM3 для CHIV (). Затем мы подтвердили некоторые из этих противовирусных факторов на уровне мРНК и белка. Однократное воздействие ВПГ и двойное воздействие ВПГ / ВИЧ увеличивало уровни экспрессии мРНК IFN-β и MX1 по сравнению с однократным воздействием ВИЧ (; Дополнительный рисунок 4), ясно показывая, что ВПГ является компонентом, ответственным за сильный противовирусный ответ.Кроме того, секреция IFN-β была выше в образцах, подвергнутых двойному воздействию, причем самые высокие уровни были вызваны HSV / HIV (). Интересно, что уровни IFN-β не коррелировали отрицательно с уровнями ВИЧ-инфекции, что позволяет предположить, что IFN-β оказывал ограниченное влияние на инфекцию даже при высоких уровнях IFN-β, наблюдаемых в ДК, подвергнутых двойному воздействию. Петля обратной связи IFN-β индуцирует активацию STAT1 и транскрипцию IFN-регулируемых генов (ISG). Наши данные ясно показали, что воздействие DC на ВИЧ индуцировало значительно более высокое фосфорилирование STAT1 через 6 часов по сравнению с CHIV, HSV / HIV и CHSV / CHIV ().Кроме того, ВИЧ и ВПГ / ВИЧ также значительно активировали несколько ISG, включая IFITM1, RSAD2 и ISG15 ().
Противовирусные факторы и пути были высокоактивированы в дендритных клетках, подвергшихся воздействию ВПГ / ВИЧ. Дендритные клетки подвергали воздействию HSV-2 или опсонизированного комплементом вируса (CHSV-2) в течение 2 часов с последующей инфекцией ВИЧ или опсонизированного комплементом ВИЧ (CHIV) в течение 22 часов. (A) Тепловая карта на основе данных последовательности РНК антивирусных факторов, значительно повышенных или понижающих регуляцию в одном или нескольких из инфекционных состояний ВИЧ, CHIV, HSV-2 / HIV или CHSV-2 / CHIV по сравнению с ложно обработанными DC ( B) Уровни экспрессии мРНК IFN-β и MXA определяли с помощью ПЦР.Данные были нормализованы к ложному набору 1. (C) Уровни секретируемого IFN-β оценивали с помощью ELISA. (D) Уровень фосфорилирования STAT1 оценивали в лизатах из DC, подвергнутых воздействию HSV-2 или CHSV-2 в течение 2 часов с последующим воздействием ВИЧ или CHIV в течение 4 часов с помощью набора фосфопротеиновых шариков. * p <0,05; ** р <0,005; *** р <0,0005. N = 5–8.
ВПГ-2 Требуется усиление ВИЧ-инфекции в ДК Интактная структура ДНК ВПГ-2
Чтобы оценить, было ли усиление ВИЧ-инфекции в ДК следствием продуктивной ВПГ-2-инфекции с полноразмерной репликацией ДНК или ее было достаточно с Для доставки вирионов интактной доступной дцДНК мы использовали ацикловир и γ-облученный инактивированный HSV-2 (51).В отсутствие репликации ДНК ВПГ-2 γ-облученный инактивированный ВПГ-2 не смог повысить уровень ВИЧ-инфекции и уровни экспрессии TNF и IFN-β (), что ясно указывает на то, что повышенная инфекция ДК ВИЧ вызывает зависит от наличия доступной и / или интактной ДНК HSV-2. Инактивация HSV-2 γ-облучением вызывает повреждение ДНК, которое может сделать вирусную ДНК менее реактивной с внутриклеточными PRR, влиять на транспорт ДНК к ядру и / или влиять на высвобождение ДНК из вирусного капсида.Мы использовали ингибитор репликации HSV-2 ацикловир, чтобы проверить, было ли присутствие интактной ДНК HSV-2 в DC достаточным для усиления ВИЧ-инфекции или требовалась ли активная репликация ДНК HSV-2. Ингибирование активной репликации ДНК HSV-2 в DC мало влияло на ВИЧ-инфекцию и IFN-β, но уменьшало ответ IFN-β, однако эти результаты не были статистически значимыми (). Чтобы дополнительно установить роль дцДНК, мы доставили HSV60, последовательность дцДНК размером 60 п.н. из HSV-1, используя DOTAP, в DC.Внутриклеточная дцДНК значительно усиливала инфекцию FHIV и CHIV по сравнению с одним только ВИЧ (). Взятые вместе, усиление ВИЧ-инфекции и повышенная продукция IFN-β в двойно экспонированных DC требовали присутствия дцДНК HSV-2 в ее естественной конфигурации, что предполагает, что активация внутриклеточных PRR, таких как сенсоры ДНК, может лежать в основе усиления HSV-2. ВИЧ-инфекции ДК.
Интактная дцДНК ВПГ необходима для усиления ВИЧ-инфекции в ДК. (A) Дендритные клетки подвергали воздействию инфекционного или гамма-облученного HSV-2 (γ-HSV-2) или опсонизированного комплементом вируса (CHSV) в течение 2 часов, затем промывали и инфицировали ВИЧ или опсонизированным комплементом ВИЧ (CHIV) в течение 22 часов. .Уровни экспрессии мРНК транскрипта gag ВИЧ-1, TNF и IFN-β определяли с помощью ПЦР. (B) ДК предварительно обрабатывали ацикловиром (20–50 мкМ) в течение 1 часа, а затем подвергали воздействию HSV или CHSV в течение 2 часов, затем промывали и инфицировали ВИЧ или CHIV в течение 22 часов с добавлением ацикловира в культуру. Уровни экспрессии генов транскрипта gag ВИЧ-1, TNF и IFN-β определяли с помощью ПЦР. (C) ДК обрабатывали HSV60, доставляемым DOTAP, или без него, а затем подвергали воздействию HSV или CHSV в течение 2 часов, затем промывали и инфицировали ВИЧ или CHIV в течение 22 часов с добавлением ацикловира в культуру.Уровни экспрессии генов транскрипта gag ВИЧ-1, TNF и IFN-β определяли с помощью ПЦР. Данные были нормализованы для ВИЧ, установленного как 1 для транскрипта gag ВИЧ-1, и ложного набора как 1 для TNF и IFN-β. * p <0,05; ** р <0,005; *** р <0,0005. N = 5–8.
Внутриклеточные PAMP, производные от HSV, являются основными факторами, участвующими в усилении инфицирования DC ВИЧ
Для оценки роли различных PAMP в усилении инфицирования DC, вызванном HSV-2, мы исследовали, являются ли TLR2, TLR3 или Активация cGAS / STING может имитировать эффекты, наблюдаемые после инфицирования HSV-2.Известно, что как TLR2, так и TLR3 вовлечены в инфекцию HSV-2 (52, 53). Стимуляция TLR2 одновременно с ВИЧ-инфекцией не влияла на экспрессию gag ВИЧ по сравнению с одним ВИЧ (). Нацеливание на TLR3 давало лишь немного увеличенную экспрессию gag ВИЧ по сравнению с резко увеличенной экспрессией gag ВИЧ в DC, подвергнутых воздействию как HSV-2, так и ВИЧ (). Хотя воспалительные реакции только на агонист TLR3 и агонист TLR3 в комбинации с ВИЧ были выше, чем воспалительные реакции на только ВИЧ, ни TNF, ни IL1-β не приблизились к уровням, наблюдаемым при двойном воздействии с HSV-2 (; данные не показаны).Увидев, что TLR2 указан как важный фактор в активации клеток, индуцированной HSV-2 (52, 54), мы исследовали эффекты ингибирования пути TRL2 антагонистическим лигандом TLR2 на ВИЧ-инфекцию. Наши результаты исключают участие передачи сигналов TLR2 в усилении ВИЧ-инфекции (дополнительный рисунок 5). Принимая во внимание, что несколько межклеточных сенсоров, участвующих в инфекции HSV, например IFI16 и cGAS, которые объединяют свои сигнальные пути путем активации STING (28, 48, 55), мы исследовали влияние активации STING на ВИЧ-инфекцию.Исследовали уровни ВИЧ-инфекции в ДК после экзогенной и эндогенной стимуляции cGAMP, активатором STING через cGAS. Экзогенно доставленный цГАМФ оказывал незначительное влияние на уровень ВИЧ-инфекции, тогда как эндогенно доставленный цГАМФ, то есть доставленный в цитозоль через DOTAP, значительно усиливал ВИЧ-инфекцию до уровней, аналогичных ДК, подвергнутым воздействию ВИЧ / ВПГ (2). Это указывает на то, что внутриклеточная ДНК, воспринимающая путь cGAS / STING, является одним из путей, участвующих в опосредованном HSV-2 усилении ВИЧ-инфекции в DC.ВИЧ-инфекция в сочетании с эндогенным цГАМФ, т.е. цГАМФ, доставленным в цитозоль, индуцировала более высокие уровни экспрессии IFN-β в ДК по сравнению с одним ВИЧ и ВИЧ в сочетании с экзогенно доставленным цГАМФ (). Более высокие уровни IFN-β, экспрессируемые ДК, подвергнутыми воздействию ВПГ / ВИЧ, по сравнению с уровнями, индуцированными ВИЧ в комбинации с эндогенным цГАМФ, указывают на то, что другие сигнальные пути / факторы могут участвовать в противовирусном ответе в дополнение к пути цГАМФ / СТИНГ. Примечательно, что высокие уровни IFN-β, вызванные двойным воздействием, не препятствовали инфицированию ВИЧ в ДК, поскольку воздействие цГАМФ и ВИЧ / ВПГ индуцировало высокие уровни gag ВИЧ ().
Нуклеотиды, происходящие из внутриклеточного HSV-2, являются основными факторами, участвующими в усилении ВИЧ-инфекции в DC. (A) Незрелые DC подвергались действию TLR2 или агонистов TLR3 по отдельности или в комбинации с ВИЧ в течение 16 часов или подвергались воздействию HSV-2 (HSV) в течение 2 часов с последующим воздействием ВИЧ в течение 16 часов. Транскрипт gag ВИЧ, экспрессию IL1-β, TNF и IFN-β оценивали с помощью qPCR. (B) cGAMP вводили напрямую или через DOTAP к DC отдельно или вместе с ВИЧ в течение 16 часов, или DC подвергали воздействию HSV в течение 2 часов с последующим контактом с ВИЧ в течение 16 часов.Транскрипт gag ВИЧ, экспрессию IL1-β, TNF и IFN-β оценивали с помощью ПЦР. Данные транскрипта gag ВИЧ были нормализованы для набора ВИЧ как 1, а данные по TNF, IL1-β, IFN-β были нормализованы для фиктивного набора как 1. * p <0,05; ** р <0,005; *** р <0,0005. N = 5–8.
Двойное воздействие ДК на ВПГ / ВИЧ, активирующее и впоследствии снижающее белковые уровни сенсоров цитозольной ДНК и факторов, участвующих в их сигнальных путях
Оценивали влияние ВПГ-2 на межклеточные сенсоры в ДК.Анализ транскриптома показал, что экспрессия цитозольной ДНК и РНК-сенсоров AIM2, MB21D1 (cGAS), IFIh2, DDX58 (RIG1) и ZBP1 (DAI) была сильно повышена, а IFI16 была немного увеличена в ДК, подвергнутых воздействию только ВИЧ, и в двойные открытые DC. Экспрессия MAVS, DDX41 и DHX9 снижалась или не зависела от различных состояний ВИЧ и ВПГ / ВИЧ (). Некоторые вирусы, включая вирусы герпеса, обладают способностью регулировать факторы врожденных и адаптивных реакций клетки путем вмешательства в передачу сигнала или деградацию белков.Например, HSV1 ICP27 подавляет экспрессию IFN типа I в макрофагах человека за счет взаимодействия с STING / TBK1 (56). Литическая реактивация вируса герпеса, связанного с саркомой Капоши, вызывает деградацию IFI16 (25), а STING разрушается протеазой вируса денге (57). Более того, стимуляция врожденных сигнальных путей, например, STING, TBK1 и IRF3, может приводить к подавлению белковой регуляции постпатогенной активации противовирусных ответов (58–60). В свете этого мы оценили уровни белка нескольких сенсоров ДНК и сигнальных молекул в ДК, подвергнутых воздействию ВИЧ, ВПГ или ВИЧ / ВПГ, соответственно, и обнаружили, что воздействие одного вируса простого герпеса и двойного вируса значительно снижает экспрессию сенсоров ДНК. , и cGAS, оба активатора пути STING, ясно указывая на то, что это, прежде всего, эффект, вызванный вирусом простого герпеса ().Не было значительных различий в эффектах, наблюдаемых с опсонизацией или без нее. Итак, затем мы исследовали экспрессию белков факторов в пути STING и не обнаружили значительного влияния на белок STING, даже если существовала четкая тенденция к снижению уровня белка STING для ВИЧ / HSV, но нижестоящий фактор TBK1 от STING был значительно снижен и наблюдалась тенденция к снижению экспрессии IRF3 в DC, подвергнутых двойному воздействию, по сравнению с одним ВИЧ (). Взятые вместе, эти результаты указывают на то, что протеолитическая деградация этих белков индуцируется в первую очередь HSV и может происходить в DC, подвергнутых воздействию HSV-2, либо как прямые эффекты вирусных белков и / или из-за вирусной активации путей с участием этих факторов, что впоследствии приводит к к их деградации (58).
ВПГ-2 ВИЧ-инфекция ДК индуцировала активацию и деградацию сенсоров цитозольной ДНК. Дендритные клетки подвергали воздействию свободного HSV-2 (HSV) или опсонизированного комплементом вируса (CHSV) в течение 2 часов, затем инфицировали ВИЧ или опсонизированным комплементом ВИЧ (CHIV) в течение 22 часов. (A) Тепловая карта по противовирусным факторам значительно повышается или понижается в одной или нескольких группах DC HIV, CHIV, HSV / HIV или CHSV / CHIV по сравнению с DC, обработанными ложно, с помощью последовательности RNA seq. (B) Уровни IFI16 и cGAS в DC в различных условиях инфицирования путем окрашивания клеток и анализа с помощью проточной цитометрии. (C) Уровни STING, TBK1 и IRF3 в DC в различных условиях инфицирования путем окрашивания клеток и анализа с помощью проточной цитометрии. *** р <0,0005. N = 5–8.
Активация пути STING была необходима для усиленной ВИЧ-инфекции DC, индуцированной HSV-2
Путь STING активируется вышестоящими датчиками ДНК, такими как cGAS и IFI16, оба реализованы в цитозольном распознавании ДНК HSV (27, 61). Кроме того, недавние данные указывают на сотрудничество между IFI16 и cGAS в активации STING (62), при этом IFI16 и cGAS играют разные роли в противовирусных ответах против герпесвирусов (63).Примечательно, что другие исследования утверждают, что cGAS является основным сенсором дцДНК, ответственным за активацию пути STING / TBK / IRF3 (29, 64). Здесь мы оценили роль cGAS / IFI16 в усилении ВИЧ-инфекции в ДК, кондиционированных HSV-2, с использованием ингибирующего олигодезоксинуклеотида A151 (65). Первоначально предполагалось, что внутриклеточный A151 ингибирует IFI16, но недавние открытия показали, что он также влияет на активность cGAS (65, 66). Ингибирующий олигодезоксинуклеотид A151 незначительно увеличивал ВИЧ-инфекцию, оцениваемую по транскрипту gag, в ранний момент времени (8 ч) в DC, подвергнутых воздействию ВИЧ и HSV / ВИЧ.Через 24 часа не было различий между обработанными A151 и необработанными DC двойного воздействия (). Интересно, что ингибирующий олигодезоксинуклеотид A151 снижал экспрессию мРНК IFN-β в условиях HSV / HIV как через 8, так и через 24 часа (). Чтобы подтвердить результат цитозоля A151, мы также протестировали эффект цитозольной доставки белка герпеса UL83, который обладает способностью вмешиваться в функции IFI16 (67) и cGAS (68), в DC, подвергнутых воздействию ВИЧ или HSV / ВИЧ. UL83 показал тот же профиль, что и A151, в отношении экспрессии транскриптов мРНК gag и IFN-β (; дополнительная фигура 6).Данные показали, что ингибирование cGAS / IFI16 ингибирующим олигодезоксинуклеотидом A151 или UL83 было недостаточно для подавления усиленной ВИЧ-инфекции, но позволило снизить, но не полностью подавить ответ IFN-β, поэтому мы не могли исключить остаточную активность. этих путей в DC. Для дальнейшего изучения роли пути STING в усилении инфекции и повышенном противовирусном ответе в двойных экспонированных DC, мы нацелены на белок STING, используя рекомбинантный белок HPV 18 E7, который, как известно, ингибирует этот путь путем связывания и блокирования STING (69, 70).Внутриклеточно доставленному E7 удалось снизить экспрессию транскрипта gag и IFN-β в двойных инфицированных DC. Снижение, однако, было незначительным для затыкания рта, но тенденцию можно было увидеть (). Затем, чтобы проверить и усилить роль STING, мы оценили роль пути STING с использованием линии клеток THP1, лишенной укуса из-за нокаута STING с помощью siRNA, и обнаружили снижение экспрессии gag в клетках, подвергшихся воздействию ВИЧ / HSV, но не получили четких доказательств этого в CHSV / CHIV (), что может быть связано с низким уровнем CR3 в THP1.Взятые вместе, наши результаты показывают, что путь STING участвует в усилении ВИЧ-инфекции в DC, подвергнутых воздействию HSV-2.
Путь STING был вовлечен в усиление ВИЧ-инфекции ДК, подвергнутых воздействию HSV-2. Ингибитор IFI16 / cGAS олигонуклеотид A151 доставлялся внутриклеточно с помощью DOTAP к DC. Затем клетки подвергали воздействию свободного HSV-2 (HSV) или опсонизированного комплементом вируса (CHSV) в течение 2 часов перед инфицированием ВИЧ или опсонизированным комплементом ВИЧ (CHIV) в течение (A) 6 часов (B) или 22 часов. (B) Ингибитор белка STING Белки E7 HPV 18 доставлялись внутриклеточно с помощью DOTAP к DC. Затем клетки подвергали воздействию свободного HSV или CHSV в течение 2 часов перед инфицированием ВИЧ или CHIV в течение 22 часов. Транскрипт gag ВИЧ и экспрессия IFN-β оценивали с помощью qPCR. (C) Клетки THP1 с нокаутом по STING или дикого типа подвергали воздействию свободного HSV-2 или опсонизированного комплементом вируса (CHSV) в течение 2 часов, затем инфицировали ВИЧ или ВИЧ, опсонизированным комплементом (CHIV), в течение 22 часов. Транскрипт gag ВИЧ и экспрессия IFN-β оценивали с помощью qPCR.Данные по gag для ВИЧ были нормализованы для свободного вируса, установленного как 1, а данные IFN-β были нормализованы для фиктивного набора, равного 1. * p <0,05; ** р <0,005; *** р <0,0005. N = 5–8.
Усиление инфицирования ВИЧ в DC, кондиционированных HSV-2, вероятно, было связано со снижением уровней регуляторных факторов ВИЧ как следствие активации пути cGAS-STING
Профили транскриптома ясно продемонстрировали комбинаторные эффекты HSV-2 и воздействие ВИЧ на многие главные регуляторы ВИЧ-инфекции в DC, включая SAMHD1, TREX, APOBEC3G, PPIA (циклофилин A) и BST2 (tetherin) ().Экспрессия генов большинства регуляторных факторов ВИЧ была увеличена в DC, инфицированных только ВИЧ или в комбинации с HSV-2, по сравнению с контролем, тогда как экспрессия SAMHD1 была снижена. Кроме того, BST2 был немного снижен в группе CHIV / CHSV. Инфекция HSV-2 может модулировать экспрессию нескольких белков (25, 71), а активация противовирусных и врожденных путей может влиять на экспрессию и уровни белка, поэтому мы исследовали влияние воздействия HSV-2 на уровни белков регуляторов ВИЧ. в РЦ.Уровни белка SAMHD1, APOBEC3G и TREX1 были значительно ниже в одинарных HSV и CHSV и в двойных DC, подвергнутых воздействию HSV / HIV и CHIV / CHSV, по сравнению с ложно обработанными клетками. ВИЧ и CHIV не оказали влияния на эти факторы или оказали минимальное влияние на эти факторы (). Затем мы оценили, зависит ли снижение регуляторных факторов ВИЧ от внутриклеточной активации пути cGAS-STING дцДНК, полученной из HSV-2, с помощью цГАМФ, который продуцируется после активации дцДНК cGAS (61, 72). ДК, подвергавшиеся как цитозольному cGAMP, так и ВИЧ, имели более низкие уровни белка как SAMHD1, так и TREX1 по сравнению с ДК, подвергавшимися только воздействию ВИЧ ().В случае APOBEC3G на экспрессию не влиял cGAMP, что указывает на более сложную активацию и / или протеолиз, индуцированный факторами, производными HSV-2, по сравнению с одним cGAMP (). Эти данные демонстрируют, что путь cGAS-STING, активируемый интактной ДНК HSV-2, способствует усилению ВИЧ-инфекции в DC, опосредованному HSV-2, за счет уменьшения количества нескольких ключевых регуляторных факторов ВИЧ.
Активация пути STING в двойных инфицированных DC снижает уровни белка нескольких факторов рестрикции ВИЧ.Дендритные клетки подвергали воздействию свободного HSV-2 или опсонизированного комплементом вируса (CHSV) в течение 2 часов, затем инфицировали ВИЧ или опсонизированным комплементом ВИЧ (CHIV) в течение 22 часов. (A) Тепловая карта из данных транскриптома РНК seq внутриклеточных сенсоров, которые значительно повышаются или понижаются в одном или нескольких из инфекционных состояний ВИЧ, CHIV, HSV / HIV или CHSV / CHIV по сравнению с ложно обработанными DC. (B) Уровни SAMHD1, ABOBEC3G и TREX1 в DC в различных условиях инфицирования оценивали путем окрашивания клеток и анализировали с помощью проточной цитометрии. (C) cGAMP вводили экзогенно или доставляли внутриклеточно через DOTAP к DC, после чего DC оставались неэкспонированными или подвергались воздействию вместе с ВИЧ в течение 16 часов. Кроме того, одна группа DC подвергалась воздействию HSV в течение 2 часов, а затем HIV или CHIV в течение 16 часов. Уровни SAMHD1, ABOBEC3G и TREX1 оценивали путем окрашивания клеток и анализировали с помощью проточной цитометрии. * p <0,05; ** р <0,005; *** р <0,0005. N = 5–8.
Обсуждение
Более глубокое понимание факторов, влияющих на ВИЧ-инфекцию, имеет большое значение, и было показано, что ранее существовавшая инфекция и / или воспаление на слизистой оболочке половых органов увеличивает риск заражения ВИЧ.В случае HSV-2 риск заражения ВИЧ-инфекцией в три раза выше (2, 9, 14). Во время первичной ВИЧ-инфекции слизистой оболочки HSV-2, как инфекционные, так и нейтрализованные вирусы, а также остатки умирающих инфицированных клеток, будут влиять на местные врожденные и адаптивные иммунные ответы. ДК, расположенные в слизистой оболочке, являются одними из первых клеток, нацеленных на ВИЧ. Здесь мы стремились установить эффекты, оказываемые кондиционированием ДК HSV-2 до заражения ВИЧ. Мы обнаружили более высокую восприимчивость и уровень продуктивной ВИЧ-инфекции в ДК, подвергшихся воздействию ВПГ-2, что зависело от структурно интактной ДНК ВПГ-2 в ДК.Примечательно, что высокие уровни продукции IFN-β, индуцированные в DC воздействием HSV-2, не смогли контролировать инфекцию, поскольку IFN-β увеличивался параллельно с увеличением инфицирования ВИЧ в DC, подвергнутых двойному воздействию. Влияние HSV-2 на репликацию ВИЧ в DC не зависело от TLR2, TLR3 или IFI16, всех трех PRR, которые, как известно, активируются вирусом герпеса (55), а скорее от активации пути cGAS-STING и последующего снижения содержания белка. уровни нескольких ключевых регуляторов ВИЧ-инфекции, т. е. TREX1, SAMHD1 и ABOBEC3G, что может быть связано с протеолитической деградацией.
ДК, локализованные в слизистой оболочке, считаются мишенью для ВИЧ-1, как это было показано на макаках, инфицированных SIV, и на модели in vitro на макаках (73–75). Первоначальное взаимодействие хозяина с вирусом в слизистой оболочке также включает активацию системы комплемента (19), и как на врожденный, так и на активированный иммунитет влияет система комплемента, и комплемент может участвовать в патогенезе хронических заболеваний (76). Вирионы, передаваемые во время половой передачи в DC, должны быть частицами, опсонизированными комплементарными антителами и / или комплементарными антителами, видимыми при контакте с семенной жидкостью и / или секрециями шейки матки (19).Результат нашего исследования с использованием ДК, полученных из моноцитов, который, как мы полагаем, является хорошей моделью для in vivo ДК ткани / слизистой оболочки, наблюдая аналогичные результаты для свободной и опсонизированной комплементом инфекции ВИЧ-1 и для инфекции ВПГ-2 в ДК слизистой оболочки шейки матки [(75) и Svanberg et al. работа в процессе]. Используемые нами ДК, происходящие из моноцитов, по-видимому, похожи на недавно определенные эпидермальные ДК CD11c +, которые являются мишенью для ВИЧ-инфекции (77). Эпидермальные CD11c + DC и клетки Лангерганса могут играть важную роль в передаче ВИЧ-1 половым путем (8, 77).Тот факт, что лишь несколько процентов ДК, подвергшихся воздействию ВИЧ и ВПГ-2, продуктивно инфицированы обоими вирусами (8), ясно указывает на то, что усиление инфицирования ДК ВИЧ требовало только воздействия вирионов ВПГ-2 или вирусной ДНК. Это создаст контекст, в котором уже существующая инфекция ВПГ-2 привлекает и предоставляет больше клеток-мишеней для ВИЧ, например, Т-лимфоциты CD4 и ДК (2, 8, 9), а также среду с микрозонами с ВПГ-2, провоспалительные цитокины и ДНК HSV-2, высвобождаемая умирающими клетками.Взятые вместе, все эти факторы создают предпосылки для ДК слизистой оболочки и делают их более восприимчивыми к продуктивной ВИЧ-инфекции.
Первоначальный анализ транскриптома продемонстрировал, что все условия инфицирования имеют различные профили экспрессии с некоторым сходством между условиями со свободными вирусами, то есть ВИЧ и ВИЧ / HSV, и между вирусами, опсонизированными комплементом, то есть CHIV и CHIV / CHSV. Многочисленные пути, участвующие в противовирусных и воспалительных ответах, были сильно затронуты в DC, инфицированных свободным ВИЧ или опсонизированным комплементом или неопсонизированным HSV / ВИЧ, тогда как опсонизированные комплементом вирусы в целом активировали меньшее количество путей.Эффективное подавление активации DC опсонизированным комплементом ВИЧ должно в первую очередь быть связано с подавлением врожденных и адаптивных путей, опосредованных сигнализацией рецептора 3 комплемента (CR3), задействованной инактивированным фрагментом C3b на вирусной поверхности (20). Предварительное кондиционирование DC с помощью HSV-2 с последующей ВИЧ-инфекцией явно усиливало активацию DC по сравнению с одной только ВИЧ-инфекцией. Это должно зависеть в первую очередь от сильной и продолжительной активации, вызываемой HSV-2, поскольку только ВИЧ вызывает слабую и более временную активацию DC.
Произошла массивная транскрипционная активация воспалительных и противовирусных факторов в ДК, подвергшихся воздействию ВИЧ / ВПГ, однако это не полностью отразилось на уровне белка без изменений или даже снижения уровней некоторых белков. Это соответствует нашим предыдущим наблюдениям для DC, инфицированных HSV-2 (18). У этого несоответствия может быть несколько причин, таких как посттранскрипционная модификация, деградация белками, производными от HSV-2 (25, 56) и / или целенаправленная протеолитическая деградация белков, участвующих в клеточных процессах, из-за вирусной активации клеточного стресса, такого как подавление факторов аутофагии и постпатогенной активации.Вирусы герпеса имеют широкий спектр стратегий, позволяющих избежать чувствительности врожденного иммунитета и повысить свою выживаемость. Обычные стратегии, используемые вирусами герпеса, заключаются в подавлении или разрушении белков, участвующих в противовирусных иммунных ответах (25, 48, 78). Некоторые белки HSV и вирусные некодирующие РНК могут модулировать экспрессию белка, например, предполагается, что белок UL41, кодируемый HSV, обладает способностью запускать быстрое и селективное отключение синтеза белка в клетке-хозяине для нескольких белков, включая виперин, и тезерин (79–82).
Было показано, что множественные PRR участвуют в клеточном распознавании и защите от герпесвирусов (55), когда мы тестировали TLR2 и TLR3, мы не обнаружили, что ни один из этих TLR не участвует в повышенной ВИЧ-инфекции, наблюдаемой в кондиционированных HSV-2 DC. . Вместо этого в недавних сообщениях указывается на активацию пути STING и его важную роль в противовирусной защите от HSV (83, 84). Поющий каскад STING может пересекать как путь TBK1 / IRF3, так и путь NFκB (85–87). Несколько сенсоров ДНК были вовлечены в активацию STING, такие как IFI16, DDX41 и cGAS, но накопленные данные ясно показали, что определение cGAS требуется для большинства типов клеток.Это верно, даже если существует сотрудничество в активации STING между IFI16 и cGAS в кератиноцитах (62), а IFI16 способствует продукции cGAS cGAMP в макрофагах человека (72). Все наши данные указывают на активацию пути cGAS STING в двойных зараженных DC. Когда дело доходит до повышенного уровня ВИЧ-инфекции, мы можем исключить участие IFI16, поскольку блокирование этого фактора не привело к снижению ВИЧ-инфекции, тогда как прямое нацеливание на STING помогло сделать то же самое. С другой стороны, мы не можем исключить участие IFI16 в распознавании двойной инфекции и взаимодействие этого сенсора с cGAS в активации противовирусных ответов IFN-β, поскольку блокирование IFI16 снижает ответ IFN-β.Важная роль IFI16 в обнаружении внутриклеточной ДНК была ранее показана в макрофагах человека посредством усиления продукции и функции цГАМФ (72).
ВИЧ-инфекция ДК жестко регулируется и ограничивается множеством противовирусных факторов, таких как SAMHD1, APOBEC3G, TRIM5α, SLFN11 и IFITMs, которые проявляют свои эффекты на разных этапах цикла репликации ВИЧ (88–90). Эти факторы контролируют или подавляют способность ВИЧ вызывать продуктивную инфекцию SAMHD1 в качестве основного подавляющего фактора ВИЧ (90, 91).SAMHD1 ингибирует ранние стадии ВИЧ-инфекции, ограничивая синтез вирусной кДНК за счет истощения дезоксинуклеотидтрифосфатов (dNTP) (90), а деградация SAMHD1 под действием VPX ВИЧ-2 приводит к усилению инфицирования DC и продукции IFN типа I (91). ВИЧ использует 3′-репарированную экзонуклеазу 1 хозяина (TREX1), наиболее распространенную 3′-5′-ДНКазу в клетках, для связывания и разрушения избыточной цитозольной дцДНК ВИЧ-1 и, таким образом, ограничения ответа IFN типа I (88). Было показано, что помимо ограничения ВИЧ-инфекции SAMHD1 и TRIM5α также участвуют в регуляции инфекции HSV-1, ограничивая репликацию ДНК (92, 93).Мы обнаружили высокую степень протеолитической деградации некоторых факторов рестрикции ВИЧ-1, включая SAMHD1, IFI16, TREX1 и APOBEC3G (88–90), в двойно экспонированных DC. Субоптимальные уровни нескольких ключевых факторов ограничения могут объяснить потерю контроля DC над ВИЧ-инфекцией и усиление продуктивной ВИЧ-инфекции в DC. Интересно, что активация пути cGAS-STING в DCs параллельно с воздействием ВИЧ индуцировала деградацию SAMHD1 и TREX1, но не APOBEC3G.Более того, внутриклеточная активация пути cGAS-STING с помощью cGAMP давала аналогичные эффекты на репликацию ВИЧ и продукцию цитокинов, как и предварительное кондиционирование с помощью HSV-2, указывая на важную роль этого пути в клеточной модуляции функций DC HSV-2. Активация пути STING в ДК, подвергшихся воздействию как ВИЧ, так и ВПГ-2, должна впоследствии привести к накоплению большего количества нуклеотидов в цитозоле из-за более низких уровней 3′-5′-экзонуклеазы TREX1 и фосфогидролазы SAMHD1, которые должны поддерживать вирусная репликация.Отсутствие деградации APOBEC3G за счет экзогенной активации cGAMP STING указывает на то, что в этом участвовали другие процессы / факторы, вызванные HSV-2, и точно определить, какие факторы еще предстоит установить, но они могут включать производные от HSV факторы или факторы, активируемые воздействием вируса. Например, открытие Марсдена и др. тот факт, что TNF, продуцируемый DC, подвергнутыми воздействию HSV-2, был вовлечен в усиление ВИЧ-инфекции сторонних DC (8), указывает на то, что TNF может быть одним из факторов, участвующих в продвижении ВИЧ-инфекции в DC.
Множество сенсоров и кофакторов цитозольной ДНК, например IFI16, cGAS, STING, TBK1 и IRF3, важных для распознавания вирусных PAMP, также были нацелены на деградацию в DC, подвергнутых двойному воздействию ВИЧ и HSV-2, но не в ВИЧ. выявили DC, демонстрируя прямое и / или косвенное участие HSV-2 в этом процессе. Уменьшение количества нескольких белков, участвующих в пути STING, например IFI16, cGAS, STING, TBK1 и IRF3, вероятно, было связано с вирусной активацией пути STING (58, 94) и / или прямой деградацией вирусных белков (95) .Чтобы проанализировать это дальше, необходимы будущие исследования фосфорилирования и локализации этих белков. Некоторые белки ДНК-вируса могут расщеплять или разрушать датчики ДНК. Например, сенсор дцДНК IFI16, который активирует ответы IFN-β и формирует инфламмасому, разрушается в клетках, инфицированных HSV-1, под действием протеасомного таргетинга ICP0 (48) и в клетках с литической саркомой Капоши-ассоциированной герпесвирусной инфекцией (25). STING и cGAS также являются обычными мишенями для белков ДНК-вирусов, которые разрушают эти факторы или мешают их передаче сигналов ниже по течению (57, 70, 96–98).В ДК, подвергнутых двойному воздействию, уровни cGAS были ниже по сравнению с неинфицированными или одиночными ВИЧ-инфицированными клетками, и это должно снизить продукцию cGAMP и активацию STING в результате клеточной регуляции и контроля этого фактора после активации ( 56). Несколько вирусов распознаются и регулируются путем STING, включая HSV-1, HSV-2 и HIV-1 (29, 56). ДНК ВИЧ-1 может распознаваться cGAS и путем STING (29), однако этому противодействует и предотвращается капсид ВИЧ-1 путем привлечения клеточных белков, например.g., циклофилин A и TRIM5α (99). HSV-1 противодействует пути STING-cGAS с помощью нескольких белков, включая ICP27, который может ингибировать экспрессию IFN типа I, воздействуя на сигнальный комплекс STING (56). Ясно, что и ВИЧ, и HSV-2 регулируются путем STING и разработали стратегии для установления инфекции путем подавления активации STING и последующих ответов IFN типа I.
Мы обнаружили и ранее показали (20), что в ДК, полученных из моноцитов, воздействие только ВИЧ-1 действительно может вызвать всплеск низкого уровня IFN-β, этот ответ временный и слабый по сравнению со многими другими вирусами. в том числе ВПГ-2 (18).Мы думаем, что это открытие согласуется с предыдущими исследованиями Harman et al. и Gringhuis et al. которые демонстрируют, что ВИЧ-1 обладает способностью с помощью нескольких механизмов вмешиваться в продукцию IFN-β и избегать противовирусной защиты хозяина (100–102). Двойное воздействие индуцировало высокий уровень противовирусного фактора IFN-β и последующих ISG, таких как MX1 и MX2, в DC через активацию ДНК HSV-2 пути STING. Высокий уровень продукции IFNβ в ДК, подвергнутых двойному свободному и комплементарному опонированию ВИЧ / ВПГ, не смог блокировать ВИЧ-инфекцию, но очевидно, что он подавлял инфекцию, поскольку ВИЧ-инфекция была дополнительно повышена в ДК, где IFN-β был подавлен. путем ингибирования ДНК-сенсора IFI16.Это продемонстрировало балансирующее действие пути STING, играющего роль как в усилении ВИЧ-инфекции, так и в контроле уровня инфекции посредством активации IFN типа I. Ранее мы установили, что ВИЧ-1 может индуцировать продукцию IFN-β в ДК зависимым от IRF1 / 7 образом, тогда как опсонизированный комплементом ВИЧ активировал IRF3, но это не давало или давало очень низкую продукцию IFN-β (20). Это, вероятно, связано с подавлением CR3, индуцированным опсонизированным комплементом вирусом, и тем, что подавление CR3 IFN-β перекрывается сильной активацией, которую HSV-2 индуцирует в DC.
Мы ясно демонстрируем, что для увеличения ВИЧ-инфекции, зависимого от HSV-2, требуется интактная дцДНК HSV-2, но не активная репликация ДНК HSV. Клеточная поддержка продуктивной ВИЧ-инфекции должна быть обусловлена регуляцией и протеолитической деградацией факторов рестрикции ВИЧ, то есть SAMHD1, APOBEC3G, IFI16 и TREX1. Механизм, ответственный за протеолитическую деградацию белков SAMHD1, TREX1 и APOBEC3G, по-видимому, облегчается активацией пути STING, который, насколько нам известно, на сегодняшний день не описан, а также действием одного или нескольких белков HSV-2. .Деградация нескольких регуляторных факторов ВИЧ, наблюдаемая при активации STING через путь cGAMP-cGAS путем добавления экзогенного cGAMP, подтверждает роль пути STING в деградации этих белков в двойных контактах с ВПГ ВИЧ. Кроме того, противовирусные и воспалительные пути, активируемые одним ВИЧ или двойным воздействием ВИЧ / ВПГ, имели различную форму в присутствии или в отсутствие молекул комплемента. Эти данные продемонстрировали, что внутриклеточное перепрограммирование HSV-2 увеличивает способность ВИЧ инфицировать DC и обеспечивает условную микросреду, благоприятную для передачи ВИЧ и возникновения инфекции.
Заявление об этике
Исследования с участием людей были рассмотрены и одобрены Шведским советом по этическому анализу Ethical Permit M173-07 и M75-08 / 2008. Письменное информированное согласие на участие не требовалось для этого исследования в соответствии с национальным законодательством и институциональными требованиями.
Вклад авторов
Исследования, разработанные EC, CS и ML. EC, RE, CS, PB, MK, JH и SN провели исследования. KE предоставил новые реагенты и аналитические инструменты.KO, EC, RE, CS, KE, ES и ML проанализировали данные. EC, KE, CS, ES и ML написали статью. ML курировал проект.
Конфликт интересов
Авторы заявляют, что исследование проводилось в отсутствие каких-либо коммерческих или финансовых отношений, которые могли бы быть истолкованы как потенциальный конфликт интересов.
Благодарности
Вычисления для RNAseq были выполнены на ресурсах, предоставленных SNIC через Уппсальский многопрофильный центр перспективных вычислений (UPPMAX) в рамках проекта b2015293.
Сноски
Финансирование. Эта работа поддержана: AI52731, Шведский исследовательский совет, Фонд исследований шведских врачей против СПИДа, Шведское агентство международного сотрудничества в области развития, SIDA SARC, VINNMER for Vinnova, Исследовательский фонд больницы университета Линчёпинга, CALF, Шведское медицинское общество и Совет медицинских исследований Юго-Восточной Швеции по ML.
Ссылки
2. Фриман Э.Е., Вайс Х.А., Глинн Дж.Р., Кросс П.Л., Уитворт Дж.А., Хейс Р.Дж.Инфекция, вызванная вирусом простого герпеса 2, увеличивает заражение ВИЧ среди мужчин и женщин: систематический обзор и метаанализ продольных исследований. СПИД. (2006) 20: 73–83. 10.1097 / 01.aids.0000198081.09337.a7 [PubMed] [CrossRef] [Google Scholar] 3. Уолд А., Линк К. Риск инфицирования вирусом иммунодефицита человека у лиц, инфицированных вирусом простого герпеса 2-го типа: метаанализ. J Infect Dis. (2002) 185: 45–52. 10.1086 / 338231 [PubMed] [CrossRef] [Google Scholar] 6. Schiffer JT, Swan DA, Corey L, Wald A. Быстрое распространение вируса и короткий период полувыведения лекарства объясняют неполную эффективность существующих противовирусных агентов, направленных против вируса простого герпеса 2.Противомикробные агенты Chemother. (2013) 57: 5820–9. 10.1128 / AAC.01114-13 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 7. Schiffer JT, Swan DA, Magaret A, Schacker TW, Wald A, Corey L. Математическое моделирование предсказывает, что повышенное выделение HSV-2 у ВИЧ-1 инфицированных людей связано с плохим иммунологическим контролем в ганглиях и слизистой оболочке гениталий. PLoS ONE. (2016) 11: e0155124. 10.1371 / journal.pone.0155124 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 8. Марсден В., Донахи Х., Бертрам К.М., Харман А.Н., Наср Н., Кеошкериан Э. и др.. Дендритные клетки, инфицированные вирусом простого герпеса 2 типа, продуцируют TNF-альфа, который усиливает экспрессию CCR5 и стимулирует продукцию ВИЧ из соседних инфицированных клеток. J Immunol. (2015) 194: 4438–45. 10.4049 / jimmunol.1401706 [PubMed] [CrossRef] [Google Scholar] 9. де Йонг М.А., де Витте Л., Тейлор М.Э., Гейтенбек ТБ. Вирус простого герпеса 2 типа повышает восприимчивость к ВИЧ-1, влияя на функцию клеток Лангерганса. J Immunol. (2010) 185: 1633–41. 10.4049 / jimmunol.07 [PubMed] [CrossRef] [Google Scholar] 10.Sartori E, Calistri A, Salata C, Del Vecchio C, Palu G, Parolin C. Инфекция вируса простого герпеса 2 типа увеличивает проникновение вируса иммунодефицита человека 1 типа в первичные макрофаги человека. Вирол Дж. (2011) 8: 166. 10.1186 / 1743-422X-8-166 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 11. Реббапрагада А., Вачихи С., Петтенгель С., Сандерджи С., Хьюибнер С., Джаоко В. и др. . Отрицательная синергия слизистой оболочки между Herpes simplex типа 2 и ВИЧ в женских половых путях. СПИД. (2007) 21: 589–98. 10.1097 / QAD.0b013e328012b896 [PubMed] [CrossRef] [Google Scholar] 12. Чжу Дж., Хладик Ф., Вудворд А., Клок А., Пэн Т., Джонстон С. и др. . Сохранение рецептор-положительных клеток ВИЧ-1 после реактивации ВПГ-2 является потенциальным механизмом увеличения заражения ВИЧ-1. Nat Med. (2009) 15: 886–92. 10.1038 / нм.2006 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 13. Мартинелли Э., Тарингер Х., Фрэнк И., Артос Дж., Пиатак М., мл., Лифсон Дж. Д. и др. . Инфекция дендритных клеток HSV-2 усиливает высокочувствительную клетку-мишень ВИЧ-1.PLoS Pathog. (2011) 7: e1002109. 10.1371 / journal.ppat.1002109 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 14. Гуд Д., Чыонг Р., Виллегас Дж., Календа Дж., Герра-Перес Н., Пятак М. и др. . Повышение экспрессии альфа4бета7, вызванное ВПГ-2, коррелирует с повышенной восприимчивостью к вагинальной инфекции SHIV (SF162P3). PLoS Pathog. (2014) 10: e1004567. 10.1371 / journal.ppat.1004567 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 15. de Jong MA, de Witte L, Oudhoff MJ, Gringhuis SI, Gallay P, Geijtenbeek TB.Агонисты TNF-альфа и TLR повышают восприимчивость к передаче ВИЧ-1 клетками Лангерганса человека ex vivo . J Clin Invest. (2008) 118: 3440–52. 10.1172 / JCI34721 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 16. Bouhlal H, Chomont N, Haeffner-Cavaillon N, Kazatchkine MD, Belec L., Hocini H. Опсонизация ВИЧ-1 комплементом спермы усиливает инфицирование эпителиальных клеток человека. J Immunol. (2002) 169: 3301–6. 10.4049 / jimmunol.169.6.3301 [PubMed] [CrossRef] [Google Scholar] 17. Reis Machado J, da Silva MV, Cavellani CL, dos Reis MA, Monteiro ML, Teixeira Vde P и др.. Иммунитет слизистых оболочек женских половых путей, ВИЧ / СПИД. Biomed Res Int. (2014) 2014: 350195. 10.1155 / 2014/350195 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 18. Crisci E, Ellegard R, Nystrom S, Rondahl E, Serrander L, Bergstrom T. и др. . Опсонизация комплемента способствует инфицированию дендритных клеток человека вирусом простого герпеса 2. J Virol. (2016) 90: 4939–50. 10.1128 / JVI.00224-16 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 19. Штойбер Х., Банки З., Вильфлингседер Д., Диерих М.П.Взаимодействие комплемента с ВИЧ на всех этапах вирусного патогенеза. Вакцина. (2008) 26: 3046–54. 10.1016 / j.vaccine.2007.12.003 [PubMed] [CrossRef] [Google Scholar] 20. Ellegard R, Crisci E, Burgener A, Sjowall C, Birse K, Westmacott G и др. . Опсонизация ВИЧ-1 комплементом приводит к снижению противовирусных и воспалительных реакций незрелых дендритных клеток через CR3. J Immunol. (2014) 193: 4590–601. 10.4049 / jimmunol.1401781 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 21. Байтай З., Спет С., Эрдей А., Диерих М.П.Передний край: продуктивная инфекция дендритных клеток ВИЧ-1 через рецептор комплемента 3 типа (CR3, CD11b / CD18). J Immunol. (2004) 173: 4775–8. 10.4049 / jimmunol.173.8.4775 [PubMed] [CrossRef] [Google Scholar] 22. Saifuddin M, Parker CJ, Peeples ME, Gorny MK, Zolla-Pazner S, Ghassemi M, et al. . Роль связанных с вирионом гликозилфосфатидилинозитол-связанных белков CD55 и CD59 в устойчивости к комплементу производных клеточной линии и первичных изолятов ВИЧ-1. J Exp Med. (1995) 182: 501–9. 10.1084 / jem.182.2.501 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 23. Bouhlal H, Chomont N, Requena M, Nasreddine N, Saidi H, Legoff J и др. . Опсонизация ВИЧ комплементом усиливает инфицирование дендритных клеток и перенос вируса в CD4 Т-клетки CR3- и DC-SIGN-зависимым образом. J Immunol. (2007) 178: 1086–95. 10.4049 / jimmunol.178.2.1086 [PubMed] [CrossRef] [Google Scholar] 24. Мельхьорсен Дж., Ринтахака Дж., Соби С., Хоран К.А., Полтаджайнен А., Остергаард Л. и др. . Раннее врожденное распознавание вируса простого герпеса в первичных макрофагах человека опосредуется MDA5 / MAVS-зависимым и независимым от MDA5 / MAVS / РНК-полимеразой III путями.J Virol. (2010) 84: 11350–8. 10.1128 / JVI.01106-10 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 25. Рой А., Датта Д., Икбал Дж., Пизано Дж., Гжиши О., Ансари М.А. и др. . Сенсор ДНК ядерного врожденного иммунитета IFI16 разрушается во время литической реактивации герпесвируса Капоши, связанного с саркомой (KSHV): роль IFI16 в поддержании латентности KSHV. J Virol. (2016) 90: 8822–41. 10.1128 / JVI.01003-16 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 26. Датта Д., Датта С., Веттил М.В., Рой А., Ансари М.А., Икбал Дж. И др.. BRCA1 регулирует опосредованное IFI16 врожденное ядерное восприятие ДНК вируса герпеса и последующую индукцию врожденного инфламмасомы и ответов интерферона-β. PLoS Pathog. (2015) 11: e1005030. 10.1371 / journal.ppat.1005030 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 27. Орзалли М.Х., Брукема Н.М., Дайнер Б.А., Хэнкс, округ Колумбия, Элде, Северная Каролина, Кристя И.М. и др. . cGAS-опосредованная стабилизация IFI16 способствует врожденной передаче сигналов во время инфицирования вирусом простого герпеса. Proc Natl Acad Sci USA. (2015) 112: E1773–81. 10.1073 / pnas.1424637112 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 28. Хансен К., Прабакаран Т., Лаустсен А., Йоргенсен С.Е., Рахбек С.Х., Дженсен С.Б. и др. . Listeria monocytogenes индуцирует экспрессию IFNβ посредством IFI16-, cGAS- и STING-зависимого пути. EMBO J. (2014) 33: 1654–66. 10.15252 / embj.201488029 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 29. Гао Д., Ву Дж., Ву Ю. Т., Ду Ф, Аро С., Ян Н. и др. . Циклическая GMP-AMP-синтаза является сенсором врожденного иммунитета ВИЧ и других ретровирусов.Наука. (2013) 341: 903–6. 10.1126 / science.1240933 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 30. Якобсен М.Р., Бак Р.О., Андерсен А., Берг Р.К., Дженсен С.Б., Тенгчуан Дж. И др. . IFI16 распознает формы ДНК лентивирусного цикла репликации и контролирует репликацию ВИЧ-1. Proc Natl Acad Sci USA. (2013) 110: E4571–80. 10.1073 / pnas.1311669110 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 31. Towers GJ, Noursadeghi M. Взаимодействие между ВИЧ-1 и автономной клеточной системой врожденного иммунитета.Клеточный микроб-хозяин. (2014) 16: 10–8. 10.1016 / j.chom.2014.06.009 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 32. Schon E, Harandi AM, Nordstrom I, Holmgren J, Eriksson K. Вакцинация дендритными клетками защищает мышей от летальности, вызванной инфекцией вируса простого генитального герпеса 2 типа. J Reprod Immunol. (2001) 50: 87–104. 10.1016 / S0165-0378 (00) 00094-2 [PubMed] [CrossRef] [Google Scholar] 33. Россио Дж. Л., Эссер М. Т., Сурьянараяна К., Шнайдер Д. К., Бесс Дж. У., мл., Васкес Г. М. и др. . Инактивация инфекционности вируса иммунодефицита человека 1 типа с сохранением конформационной и функциональной целостности поверхностных белков вириона.J Virol. (1998) 72: 7992–8001. [Бесплатная статья PMC] [PubMed] [Google Scholar] 34. Куч О., Бенвенист Э. Н., Шоу Г. М., Леви Д. Н.. Прямой и количественный одноклеточный анализ реактивации вируса иммунодефицита человека 1 типа с латентного периода. J Virol. (2002) 76: 8776–86. 10.1128 / JVI.76.17.8776-8786.2002 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 35. Леви Д. Н., Альдрованди Г. М., Куч О., Шоу Г. М.. Динамика рекомбинации ВИЧ-1 в его естественных клетках-мишенях. Proc Natl Acad Sci USA. (2004) 101: 4204–9. 10.1073 / pnas.0306764101 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 36. Tjomsland V, Ellegard R, Che K, Hinkula J, Lifson JD, Larsson M. Комплементарная опсонизация ВИЧ-1 усиливает захват дендритными клетками и включает в себя семейства рецепторов эндоцитарного лектина и интегрина. PLoS ONE. (2011) 6: e23542. 10.1371 / journal.pone.0023542 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 37. Вандесомпеле Дж., Де Претер К., Паттин Ф., Поппе Б., Ван Рой Н., Де Паэпе А. и др. . Точная нормализация количественных данных ОТ-ПЦР в реальном времени путем геометрического усреднения нескольких генов внутреннего контроля.Genome Biol. (2002) 3: 0034. 10.1186 / gb-2002-3-7-research0034 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 39. Бабрахам-биоинформатика FastQC: инструмент контроля качества для данных последовательности с высокой пропускной способностью. Кембридж: Бабрахам-Биоинформатика; (2017). [Google Scholar] 41. Добин А., Дэвис К.А., Шлезингер Ф., Дренкоу Дж., Залески С., Джа С. и др. . STAR: сверхбыстрый универсальный выравниватель RNA-seq. Биоинформатика. (2013) 29: 15–21. 10.1093 / bioinformatics / bts635 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 42.Liao Y, Smyth GK, Shi W. featureCounts: эффективная программа общего назначения для сопоставления считываний последовательностей с геномными особенностями. Биоинформатика. (2014) 30: 923–30. 10.1093 / bioinformatics / btt656 [PubMed] [CrossRef] [Google Scholar] 43. Лав М.И., Хубер В., Андерс С. Умеренная оценка кратного изменения и дисперсии данных РНК-seq с помощью DESeq2. Genome Biol. (2014) 15: 550. 10.1186 / s13059-014-0550-8 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 44. Goldwich A, Prechtel AT, Muhl-Zurbes P, Pangratz NM, Stossel H, Romani N, et al.. Вирус простого герпеса типа I (HSV-1) реплицируется в зрелых дендритных клетках, но может передаваться только в зависимости от контакта между клетками. J Leukoc Biol. (2011) 89: 973–9. 10.1189 / jlb.0310180 [PubMed] [CrossRef] [Google Scholar] 45. Abaitua F, Hollinshead M, Bolstad M, Crump CM, O’Hare P. Сигнал ядерной локализации в герпесвирусном белке VP1-2 важен для инфицирования через путь капсида к ядерной поре. J Virol. (2012) 86: 8998–9014. 10.1128 / JVI.01209-12 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 46.Mikloska Z, Bosnjak L, Cunningham AL. Незрелые дендритные клетки, полученные из моноцитов, продуктивно инфицированы вирусом простого герпеса типа 1. J Virol. (2001) 75: 5958–64. 10.1128 / JVI.75.13.5958-5964.2001 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 47. Гао Г., Луо Х. Путь убиквитин-протеасома при вирусных инфекциях. Может J Physiol Pharmacol. (2006) 84: 5–14. 10.1139 / y05-144 [PubMed] [CrossRef] [Google Scholar] 48. Орзалли MH, DeLuca NA, Knipe DM. Ядерная индукция IFI16 передачи сигналов IRF-3 во время герпесвирусной инфекции и деградация IFI16 вирусным белком ICP0.Proc Natl Acad Sci USA. (2012) 109: E3008–17. 10.1073 / pnas.1211302109 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 49. Лю Ю., Ли М., Чжан Д., Чжан М., Ху К. Гликопротеин gD HSV-2 нацелен на CC домен тетерина и способствует его деградации через лизосомный путь. Вирол Дж. (2016) 13: 154. 10.1186 / s12985-016-0610-7 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 50. Ushijima Y, Luo C, Kamakura M, Goshima F, Kimura H, Nishiyama Y. Вирус простого герпеса UL56 взаимодействует с зудом убиквитинлигазы семейства Nedd4 и регулирует его.Вирол Дж. (2010) 7: 179. 10.1186 / 1743-422X-7-179 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 51. Стефаниду М., Рамос И., Мас Касулло В., Трепаниер Дж. Б., Розенбаум С., Фернандес-Сесма А. и др. . Вирус простого герпеса 2 (HSV-2) предотвращает созревание дендритных клеток, индуцирует апоптоз и запускает высвобождение провоспалительных цитокинов: потенциальные связи с синергией HSV-HIV. J Virol. (2013) 87: 1443–53. 10.1128 / JVI.01302-12 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 52. Сато А., Линехан М.М., Ивасаки А.Двойное распознавание вирусов простого герпеса TLR2 и TLR9 в дендритных клетках. Proc Natl Acad Sci USA. (2006) 103: 17343–8. 10.1073 / pnas.0605102103 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 53. Свенссон А., Тунбэк П., Нордстром И., Падюков Л., Эрикссон К. Полиморфизмы в Toll-подобном рецепторе 3 придают естественную устойчивость к инфекции вируса простого герпеса 2 типа. J Gen Virol. (2012) 93: 1717–24. 10.1099 / vir.0.042572-0 [PubMed] [CrossRef] [Google Scholar] 54. Триантафилу К., Эриилмазлар Д., Триантафилу М.Активация, вызванная вирусом простого герпеса 2, в вагинальных клетках включает Toll-подобные рецепторы 2 и 9 и ДНК-сенсоры DAI и IFI16. Am J Obstet Gynecol. (2014) 210: 122.e1–10. 10.1016 / j.ajog.2013.09.034 [PubMed] [CrossRef] [Google Scholar] 56. Christensen MH, Jensen SB, Miettinen JJ, Luecke S, Prabakaran T., Reinert LS, et al. . HSV-1 ICP27 нацелен на активированный TBK1 сигналсом STING, чтобы ингибировать индуцированную вирусом экспрессию IFN типа I. EMBO J. (2016) 35: 1385–99. 10.15252 / embj.201593458 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 57.Агирре С., Маэстре А.М., Пагни С., Патель Дж. Р., Сэвидж Т., Гутман Д. и др. . DENV подавляет продукцию IFN типа I в инфицированных клетках путем расщепления STING человека. PLoS Pathog. (2012) 8: e1002934. 10.1371 / journal.ppat.1002934 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 58. Конно Х., Конно К., Парикмахер Г.Н. Циклические динуклеотиды запускают фосфорилирование ULK1 (ATG1) STING для предотвращения устойчивой передачи сигналов врожденного иммунитета. Клетка. (2013) 155: 688–98. 10.1016 / j.cell.2013.09.049 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 59.Хиггс Р., Ни Габанн Дж., Бен Ларби Н., Брин Е.П., Фицджеральд К.А., Джеффрис Калифорния. Убиквитинлигаза E3 Ro52 отрицательно регулирует постпатогенное распознавание продукции IFN-бета посредством полиубиквитин-опосредованной деградации IRF3. J Immunol. (2008) 181: 1780–6. 10.4049 / jimmunol.181.3.1780 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 60. Цуй Дж., Ли Й, Чжу Л., Лю Д., Сунъян З., Ван Х. Ю. и др. . NLRP4 негативно регулирует передачу сигналов интерферона I типа, нацеливаясь на киназу TBK1 для деградации через убиквитинлигазу DTX4.Nat Immunol. (2012) 13: 387–95. 10.1038 / ni.2239 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 61. Тао Дж, Чжоу X, Цзян З. cGAS-cGAMP-STING: три мушкетера цитозольной ДНК, чувствительной и сигнальной. IUBMB Life. (2016) 68: 858–70. 10.1002 / iub.1566 [PubMed] [CrossRef] [Google Scholar] 62. Almine JF, O’Hare CA, Dunphy G, Haga IR, Naik RJ, Atrih A и др. . IFI16 и cGAS взаимодействуют в активации STING во время зондирования ДНК в кератиноцитах человека. Nat Commun. (2017) 8: 14392. 10.1038 / ncomms14392 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 63.Diner BA, Lum KK, Toettcher JE, Cristea IM. Сенсоры вирусной ДНК IFI16 и циклическая GMP-AMP-синтаза обладают различными функциями в регулировании экспрессии вирусных генов, иммунной защиты и апоптотических реакций во время герпесвирусной инфекции. MBio. (2016) 7: e01553–16. 10.1128 / mBio.01553-16 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 64. Luecke S, Holleufer A, Christensen MH, Jonsson KL, Boni GA, Sorensen LK и др. . cGAS активируется ДНК в зависимости от длины. EMBO Rep. (2017) 18: 1707–15.10.15252 / embr.201744017 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 65. Камински Дж. Дж., Шаттген С.А., Ценг Т.С., Боде С., Клинман Д.М., Фицджеральд К.А. Синтетические олигодезоксинуклеотиды, содержащие супрессивные мотивы TTAGGG, ингибируют активацию инфламмасомы AIM2. J Immunol. (2013) 191: 3876–83. 10.4049 / jimmunol.1300530 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 66. Штейнхаген Ф., Циллингер Т., Пойкерт К., Фокс М., Тудиум М, Барчет В. и др. . Супрессивные олигодезоксинуклеотиды, содержащие мотивы TTAGGG, ингибируют активацию cGAS в моноцитах человека.Eur J Immunol. (2018) 48: 605–11. 10.1002 / eji.201747338 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 67. Ли Т, Чен Дж, Кристя И.М. Тегументный белок цитомегаловируса человека pUL83 ингибирует опосредованное IFI16 определение ДНК для уклонения от иммунитета. Клеточный микроб-хозяин. (2013) 14: 591–9. 10.1016 / j.chom.2013.10.007 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 68. Биолатти М., Делль’Осте В., Паутассо С., Гуглиси Ф., фон Эйнем Дж., Крапп С. и др. . Тегументный белок цитомегаловируса человека pp65 (pUL83) подавляет продукцию интерферона I типа, инактивируя ДНК-сенсор cGAS, не влияя на STING.J Virol. (2018) 92: e01774–17. 10.1128 / JVI.01774-17 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 69. Чен К., Сун Л., Чен З. Дж. Регуляция и функция пути cGAS-STING зондирования цитозольной ДНК. Nat Immunol. (2016) 17: 1142–9. 10.1038 / ni.3558 [PubMed] [CrossRef] [Google Scholar] 70. Лау Л., Грей Е.Е., Брюнетка Р.Л., Стетсон ДБ. Онкогены ДНК опухолевого вируса противодействуют пути обнаружения ДНК cGAS-STING. Наука. (2015) 350: 568–71. 10.1126 / science.aab3291 [PubMed] [CrossRef] [Google Scholar] 71.Антониос Д., Руссо П., Ларанге А., Кердин-Ромер С., Палларди М. Механизмы синтеза ИЛ-12 дендритными клетками человека, обработанными химическим сенсибилизатором NiSO4. J Immunol. (2010) 185: 89–98. 10.4049 / jimmunol.02 [PubMed] [CrossRef] [Google Scholar] 72. Йонссон К.Л., Лаустсен А., Крапп С., Шкипер К.А., Тавачельвам К., Хоттер Д. и др. . IFI16 необходим для определения ДНК в макрофагах человека, способствуя выработке и функции цГАМФ. Nat Commun. (2017) 8: 14391. 10.1038 / ncomms14391 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 73.Ху К., Фрэнк И., Уильямс В., Сантос Дж. Дж., Уоттс П., Гриффин Дж. Э. и др. . Блокада рецепторов прикрепления и слияния подавляет инфицирование ВИЧ-1 тканью шейки матки человека. J Exp Med. (2004) 199: 1065–75. 10.1084 / jem.20022212 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 74. Zhou Z, Barry de Longchamps N, Schmitt A, Zerbib M, Vacher-Lavenu MC, Bomsel M, et al. . Эффективное проникновение ВИЧ-1 во внутреннюю крайнюю плоть опосредуется повышенным уровнем CCL5 / RANTES, который привлекает Т-клетки и способствует образованию конъюгата с клетками Лангерганса.PLoS Pathog. (2011) 7: e1002100. 10.1371 / journal.ppat.1002100 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 75. Tjomsland V, Ellegard R, Kjolhede P, Wodlin NB, Hinkula J, Lifson JD и др. . Блокирование интегринов подавляет инфицирование ВИЧ-1 иммунных клеток слизистой оболочки шейки матки человека свободными и опсонизированными комплементом вирионами. Eur J Immunol. (2013) 43: 2361–72. 10.1002 / eji.201243257 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 76. Рейс Е.С., Мастеллос, округ Колумбия, Хаджишенгаллис Г., Ламбрис Дж. Д.. Новое понимание иммунных функций комплемента.Nat Rev Immunol. (2019) 19: 503–16. 10.1038 / s41577-019-0168-x [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 77. Бертрам К.М., Боттинг Р.А., Бахарлу Х., Родс Дж. У., Рана Х., Грэм Дж. Д. и др. . Идентификация эпидермальных дендритных клеток человека, передающих CD11c (+). Nat Commun. (2019) 10: 2759. 10.1038 / s41467-019-10697-w [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 78. Кучет-Лоренко Д., Андерсон Г., Слоан Э, Орр А., Эверетт Р. Д.. Вирусная убиквитинлигаза ICP0 не является ни достаточной, ни необходимой для деградации сенсора клеточной ДНК IFI16 во время инфицирования вирусом простого герпеса 1.J Virol. (2013) 87: 13422–32. 10.1128 / JVI.02474-13 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 79. Эсклатин А., Таддео Б., Ройзман Б. Белок UL41 вируса простого герпеса опосредует селективную стабилизацию или деградацию клеточных мРНК. Proc Natl Acad Sci USA. (2004) 101: 18165–70. 10.1073 / pnas.0408272102 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 80. Шен Г., Ван К., Ван С., Цай М., Ли М.Л., Чжэн С. Вирус простого герпеса 1 противодействует виперину через свой вирионный белок-хозяин UL41.J Virol. (2014) 88: 12163–6. 10.1128 / JVI.01380-14 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 81. Зеннер Х.Л., Маурисио Р., Бантинг Дж., Крамп С.М. Вирус простого герпеса 1 противодействует ограничению тетерина за счет своей активности отключения вириона-хозяина. J Virol. (2013) 87: 13115–23. 10.1128 / JVI.02167-13 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 82. Маккаскилл Дж. Л., Рессел С., Альбер А., Редфорд Дж., Пауэр Ю. Ф., Шварце Дж. И др. . Широкий спектр ингибирования респираторной вирусной инфекции с помощью имитаторов микроРНК, направленных на передачу сигналов p38 MAPK.Мол тер нуклеиновых кислот. (2017) 7: 256–66. 10.1016 / j.omtn.2017.03.008 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 83. Deschamps T, Kalamvoki M. Внеклеточные везикулы, высвобождаемые инфицированными клетками вируса простого герпеса 1, блокируют репликацию вируса в реципиентных клетках зависимым от STING образом. J Virol. (2018) 92: e01102–18. 10.1128 / JVI.01102-18 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 84. Skouboe MK, Knudsen A, Reinert LS, Boularan C, Lioux T, Perouzel E, et al. . Агонисты СТИНГА обеспечивают перекрестный обмен антивирусами между клетками человека и обеспечивают защиту от генитального герпеса у мышей.PLoS Pathog. (2018) 14: e1006976. 10.1371 / journal.ppat.1006976 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 85. Fang R, Wang C, Jiang Q, Lv M, Gao P, Yu X и др. . NEMO-IKKβ необходимы для активации IRF3 и NF-κB в пути cGAS-STING. J Immunol. (2017) 199: 3222–33. 10.4049 / jimmunol.1700699 [PubMed] [CrossRef] [Google Scholar] 86. Эйб Т., Парикмахер Г.Н. Опосредованная цитозольной ДНК, STING-зависимая индукция провоспалительного гена требует канонической активации NF-kappaB посредством TBK1. J Virol.(2014) 88: 5328–41. 10.1128 / JVI.00037-14 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 87. Като К., Исии Р., Гото Е., Иситани Р., Токунага Ф., Нуреки О. Структурный и функциональный анализ ДНК-зондирования и иммунной активации человеческим cGAS. PLoS ONE. (2013) 8: e76983. 10.1371 / journal.pone.0076983 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 88. Yan N, Regalado-Magdos AD, Stiggelbout B, Lee-Kirsch MA, Lieberman J. Цитозольная экзонуклеаза TREX1 ингибирует врожденный иммунный ответ на вирус иммунодефицита человека 1 типа.Nat Immunol. (2010) 11: 1005–13. 10.1038 / ni.1941 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 89. Альтфельд М., Гейл М., младший. Врожденный иммунитет против инфекции ВИЧ-1. Nat Immunol. (2015) 16: 554–62. 10.1038 / ni.3157 [PubMed] [CrossRef] [Google Scholar] 90. Laguette N, Sobhian B, Casartelli N, Ringeard M, Chable-Bessia C, Segeral E, et al. . SAMHD1 представляет собой специфический для дендритных и миелоидных клеток фактор рестрикции ВИЧ-1, которому противодействует Vpx. Природа. (2011) 474: 654–7. 10.1038 / nature10117 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 91.Hertoghs N, van der Aar AM, Setiawan LC, Kootstra NA, Gringhuis SI, Geijtenbeek TB. Деградация SAMHD1 усиливает активное подавление созревания дендритных клеток ВИЧ-1. J Immunol. (2015) 194: 4431–7. 10.4049 / jimmunol.1403016 [PubMed] [CrossRef] [Google Scholar] 92. Kim ET, White TE, Brandariz-Nunez A, Diaz-Griffero F, Weitzman MD. SAMHD1 ограничивает вирус простого герпеса 1 в макрофагах, ограничивая репликацию ДНК. J Virol. (2013) 87: 12949–56. 10.1128 / JVI.02291-13 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 93.Reszka N, Zhou C, Song B, Sodroski JG, Knipe DM. Белки обезьяны TRIM5α снижают репликацию вируса простого герпеса. Вирусология. (2010) 398: 243–50. 10.1016 / j.virol.2009.11.041 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 94. Гонугунта В.К., Сакаи Т., Покатаев В., Ян К., Ву Дж., Доббс Н. и др. . Опосредованная торговлей людьми деградация STING требует сортировки до подкисленных эндолизосом и может быть нацелена на усиление противоопухолевого ответа. Cell Rep. (2017) 21: 3234–42. 10.1016 / j.celrep.2017.11.061 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 96.Агирре С., Лутра П., Санчес-Апарисио М.Т., Маэстре А.М., Патель Дж., Ламот Ф. и др. . Белок NS2B вируса денге нацелен на деградацию cGAS и предотвращает зондирование митохондриальной ДНК во время инфекции. Nat Microbiol. (2017) 2: 17037. 10.1038 / nmicrobiol.2017.37 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 97. Sun C, Schattgen SA, Pisitkun P, Jorgensen JP, Hilterbrand AT, Wang LJ, et al. . Уклонение от врожденного цитозольного восприятия ДНК гамма-герпесвирусом способствует установлению скрытой инфекции.J Immunol. (2015) 194: 1819–31. 10.4049 / jimmunol.1402495 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 98. Wu JJ, Li W, Shao Y, Avey D, Fu B, Gillen J и др. . Ингибирование зондирования ДНК cGAS белком вириона герпесвируса. Клеточный микроб-хозяин. (2015) 18: 333–44. 10.1016 / j.chom.2015.07.015 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 99. Rasaiyaah J, Tan CP, Fletcher AJ, Price AJ, Blondeau C, Hilditch L, et al. . ВИЧ-1 уклоняется от распознавания врожденным иммунитетом за счет набора специфических кофакторов.Природа. (2013) 503: 402–5. 10.1038 / nature12769 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 100. Харман А.Н., Лай Дж., Турвилл С., Самараджива С., Грей Л., Марсден В. и др. . ВИЧ-инфекция дендритных клеток нарушает путь индукции IFN через IRF-1 и подавляет продукцию IFN типа 1. Кровь. (2011) 118: 298–308. 10.1182 / blood-2010-07-297721 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 101. Харман А.Н., Наср Н., Фитхам А., Галоян А., Альшехри А.А., Рамбуквелле Д. и др. . ВИЧ блокирует индукцию интерферона в дендритных клетках и макрофагах человека за счет нарушения регуляции TBK1.J Virol. (2015) 89: 6575–84. 10.1128 / JVI.00889-15 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 102. Gringhuis SI, Hertoghs N, Kaptein TM, Zijlstra-Willems EM, Sarrami-Forooshani R, Sprokholt JK, et al. ВИЧ-1 блокирует сигнальный адаптер MAVS, чтобы избежать противовирусной защиты хозяина после восприятия абортивной РНК ВИЧ-1 хеликазой DDX3 хозяина. Nat Immunol. (2017) 18: 225–35. 10.1038 / ni.3647 [PubMed] [CrossRef] [Google Scholar]шестнадцатеричных, RGB и CMYK цветовых кодов
оттенков синего :: шестнадцатеричные, RGB и CMYK цветовые коды- Дом
Наборы цветов
Цветовые схемы
Цвета социальных сетей
Цвета бренда
Голубой
Hex: # 89cff0RGB: 137, 240, 207
CMYK: 43, 14, 0, 6
HSL: 199, 77%, 74%
HSV: 199, 43%, 94%
Beau Blue
Hex: # bcd4e6RGB: 188, 230, 212
CMYK: 18, 8, 0, 10
HSL: 206, 46%, 82%
HSV: 206, 18%, 90%
Пузыри
Hex: # e7feffRGB: 231, 255, 254
CMYK: 9, 0, 0, 0
HSL: 183, 100%, 95%
HSV: 183, 9%, 100%
CMYK Синий
Hex: # 333399RGB: 51, 153, 51
CMYK: 67, 67, 0, 40
HSL: 240, 50%, 40%
HSV: 240, 67%, 60%
Crayola Синий
Hex: # 1f75feRGB: 31, 254, 117
CMYK: 88, 54, 0, 0
HSL: 217, 99%, 56%
HSV: 217, 88%, 100%
Темно-синий
Hex: # 00008bRGB: 0, 139, 0
CMYK: 100, 100, 0, 45
HSL: 240, 100%, 27%
HSV: 240, 100%, 55%
Делфтский синий
Hex: # 1f305eRGB: 31, 94, 48
CMYK: 67, 49, 0, 63
HSL: 224, 50%, 25%
HSV: 224, 67%, 37%
Утка синяя
Hex: # 007791RGB: 0, 145, 119
CMYK: 100, 18, 0, 43
HSL: 191, 100%, 28%
HSV: 191, 100%, 57%
Голубой лед
Hex: # 99ffffRGB: 153, 255, 255
CMYK: 40, 0, 0, 0
HSL: 180, 100%, 80%
HSV: 180, 40%, 100%
Голубая Либерти
Hex: # 545aa7RGB: 84, 167, 90
CMYK: 50, 46, 0, 35
HSL: 236, 33%, 49%
HSV: 236, 50%, 65%
Голубой
Hex: # add8e6RGB: 173, 230, 216
CMYK: 25, 6, 0, 10
HSL: 195, 53%, 79%
HSV: 195, 25%, 90%
Средний синий
Hex: # 0000cdRGB: 0, 205, 0
CMYK: 100, 100, 0, 20
HSL: 240, 100%, 40%
HSV: 240, 100%, 80%
Утренний синий
Hex: # 8da399RGB: 141, 153, 163
CMYK: 13, 0, 6, 36
HSL: 153, 11%, 60%
HSV: 153, 13%, 64%
Манселл синий
Hex: # 0093afRGB: 0, 175, 147
CMYK: 100, 16, 0, 31
HSL: 190, 100%, 34%
HSV: 190, 100%, 69%
Темно-синий
Hex: # 000080RGB: 0, 128, 0
CMYK: 100, 100, 0, 50
HSL: 240, 100%, 25%
HSV: 240, 100%, 50%
NCS Синий
Hex: # 0087bdRGB: 0, 189, 135
CMYK: 100, 29, 0, 26
HSL: 197, 100%, 37%
HSV: 197, 100%, 74%
Неоново-синий
Hex: # 4666ffRGB: 70, 255, 102
CMYK: 73, 60, 0, 0
HSL: 230, 100%, 64%
HSV: 230, 73%, 100%
Синий Pantone
Hex: # 0018a8RGB: 0, 168, 24
CMYK: 100, 86, 0, 34
HSL: 231, 100%, 33%
HSV: 231, 100%, 66%
Барвинок
Hex: #ccccffRGB: 204, 255, 204
CMYK: 20, 20, 0, 0
HSL: 240, 100%, 90%
HSV: 240, 20%, 100%
Пикоти Синий
Hex: # 2e2787RGB: 46, 135, 39
CMYK: 66, 71, 0, 47
HSL: 244, 55%, 34%
HSV: 244, 71%, 53%
Пудрово-синий
Hex: # b0e0e6RGB: 176, 230, 224
CMYK: 23, 3, 0, 10
HSL: 187, 52%, 80%
HSV: 187, 23%, 90%
RGB Синий
Hex: # 0000ffRGB: 0, 255, 0
CMYK: 100, 100, 0, 0
HSL: 240, 100%, 50%
HSV: 240, 100%, 100%
Красно-синий
Hex: # 76abdfRGB: 118, 223, 171
CMYK: 47, 23, 0, 13
HSL: 210, 62%, 67%
HSV: 210, 47%, 87%
Сапфир
Hex: # 082567RGB: 8, 103, 37
CMYK: 92, 64, 0, 60
HSL: 222, 86%, 22%
HSV: 222, 92%, 40%
Испанский синий
Hex: # 0070bbRGB: 0, 187, 112
CMYK: 100, 40, 0, 27
HSL: 204, 100%, 37%
HSV: 204, 100%, 73%
бирюзовый
Hex: # 008080RGB: 0, 128, 128
CMYK: 100, 0, 0, 50
HSL: 180, 100%, 25%
HSV: 180, 100%, 50%
Бирюзово-голубой
Hex: # 367588RGB: 54, 136, 117
CMYK: 60, 14, 0, 47
HSL: 194, 43%, 37%
HSV: 194, 60%, 53%
Ультрамарин
Hex: # 4000ffRGB: 64, 255, 0
CMYK: 75, 100, 0, 0
HSL: 255, 100%, 50%
HSV: 255, 100%, 100%
Уранский синий
Hex: # afdbf5RGB: 175, 245, 219
CMYK: 29, 11, 0, 4
HSL: 202, 78%, 82%
HSV: 202, 29%, 96%
Многофункциональный очаровательный цветок Ароматные цветы Семена дерева глицинии CHSV
Главнаяjimrobinson2019-02-07T12: 23: 03 + 00: 00Многофункциональный очаровательный цветок Ароматные цветы Семена дерева глицинии CHSV
Портфель через плечо из натуральной кожи 81stgeneration Винтажная сумка для ноутбука: Clothing.Лазерное травление — более дешевый процесс, который нагревает стекло и оставляет на поверхности крошечные трещины, которые проявляются в виде дефектов дизайна. С хорошей амортизацией / амортизацией, сверхпрочным трикотажным материалом из полиэстера, покупайте эластичные повседневные рубашки на пуговицах с длинным рукавом и другие повседневные рубашки на пуговицах yibiyuan в, Наш широкий выбор элегантен для бесплатной доставки и бесплатного возврата. Подходит для всех типов плит, кроме индукционных. Впервые появившийся в комиксах DC в 38 году, этот сантехнический фитинг с концентрическим переходником с полированным сварным швом соответствует санитарным стандартам 3-A № 63-03 для санитарных фитингов (номер разрешения 61) и подходит для использования в пищевых продуктах.Сделано с помощью техники сублимации и профессионального термопресса. составляет примерно 10 рабочих дней, включая отслеживание и страховку. Инструкции по стирке: машинная стирка в деликатном режиме. — Не нужно ждать — у вас будет доступ к вашему товару сразу после покупки. ● Ремень для переноски из хлопковой тесьмы шириной 1 дюйм. Все фотографии сделаны при естественном рассеянном дневном свете. Белый хлопок Вязание крючком Занавеска для окон ручной работы, *** Я показываю 4 возможных варианта тесьмы на фотографиях — если у вас есть предпочтения. Быстросохнущая эластичная ткань для удобной посадки.Наименование товара; простая трехслойная железная ваза для фруктов. женские брюки-универсал на спортивной подкладке для подъема бедер. гибко регулируется двумя подвижными шарнирами. Купить мужские кроссовки для тренинга ASICS Gel-Pulse 11 ✓ бесплатная доставка ✓ бесплатный возврат соответствующих заказов. Моющийся и герметичный (темно-синий L): товары для домашних животных, два вентиляционных отверстия предотвращают конденсацию.
Многофункциональный очаровательный цветок Ароматные цветы Семена дерева глицинии CHSV
Убежище Департамента гражданской обороны Олово-металлический знак Бомбоубежище Вторая мировая война, холодная война, HEPA-фильтр Fellowes Aeramax 300 с 4 сменными угольными фильтрами от GoodVac.2-ярусная 8-дюймовая бамбуковая пароварка, китайская корзина для рисования дим-сам, набор для рисовой пасты с крышкой!, 12×18 12 дюймов x18 дюймов, оптовая продажа, Франция, Франция, Франция и Канада, канадский флаг палки, один из самых супер-острых сортов перца чили. Бронзовый тон Париж Статуэтка Эйфелевой башни Статуя из сплава Модель Декор 5CM P0CACYC. Хромированный настенный поворотный рычаг с двойной ручкой Поворотный излив Стандартный кухонный кран. Жаккардовое кухонное полотенце Garnier Thiebaut для кухни Французское MILLE TILES Многоцветный $ 23,50, Морской узел веревочного узла Место для визиток Место назначения Пляж Свадебные сувениры, 100шт сочные растения семена кактуса домашний офис балкон бонсай орнамент декоры.Забавная наклейка на машину для ноутбука СОЛНЕЧНЫЙ СВЕТ, СМЕШАННЫЙ С МАЛЕНЬКИМ УРАГАНОМ. Виниловая наклейка, фильтры. Быстрый фильтр для воды увлажнителя A10PR № 10 для Aprilaire 550 10-PACK, 1 шт. Спиральный нож Tornado для картофеля Ручной слайсер Изготовление Twist Shredder Kitchen. 3 » / 6 » Многоразовые резиновые стяжки Magic Rubber Twist Tie Cable Wire Gear Tie 1шт / 5шт / 10шт. Крапива Травяной аромат благовоний Magikal Potion Ritual Wicca Pagan Goth Altar. ДАЙТЕ МНЕ ТЕРПЕНИЕ РАЗБРАТЬСЯ С МОИМИ БЛАГОСЛОВЕНИЯМИ Наклейка на стену Цитата Слова Буквы, OEM Рем Рууд Ричмонд Vanguard Honeywell Водонагреватель LP газовый клапан AP14672-2.
java — Каков диапазон значений HSV для КОРИЧНЕВОГО цвета в openCV?
Вы можете ознакомиться с моим ответом здесь, в котором подробно рассказывается о том, как построить / просмотреть диапазон различных значений HSV.
Вы также можете посетить внешние сайты, которые позволяют вам поиграть со значениями, чтобы получить представление о цветовом пространстве. В HSV коричневые цвета соответствуют более высокому уровню насыщенности (более низкие значения становятся более серыми) и средним уровням значений (низкий — черный, высокий — белый), а оттенок находится между (примерно) 20 градусами для красноватого кирпичного цвета и 40 градусами для песочный цвет.Но в OpenCV степени оттенка делятся на два, чтобы они соответствовали 255, так что это больше похоже на 10-20 для значений оттенка.
Используя код из связанного ответа, я создал этот gif из диапазона коричневых значений от [10, 100, 20] до [20, 255, 200] в HSV:
Кажется, это охватывает большинство идей коричневого, но вы можете поиграть со значениями, если хотите, чтобы появились более темные, светлые, серые и т. Д.
Еще один способ поиграться с поиском хороших пороговых значений — создать программу, которая позволит вам опробовать ее.Например, я создал инструмент на Python и OpenCV, похожий на селектор волшебной палочки в фотошопе, который позволяет вам определять порог, щелкать пиксель, и он будет выбирать пиксели аналогичного цвета, связанные с тем местом, где вы щелкнули. Этот инструмент распечатает самые высокие и самые низкие значения цвета, а также распечатает средний цвет и стандартное отклонение для лучшего контроля значений порога, аналогичных выбранному вами цвету. Вы можете увидеть, что здесь используется небольшой фрагмент gif .
Еще один способ поэкспериментировать со значениями — создать панели треков с OpenCV, дающие все минимальные и максимальные значения порога, и обновление экрана с изображением порога каждый раз, когда вы меняете значение.Я создал инструмент для этого также на Python и OpenCV. Вы можете увидеть, что здесь используется небольшой фрагмент gif .
Если вы внимательно изучите код обоих этих проектов, вы сможете получить представление о том, как создавать похожие программы, чтобы экспериментировать со значениями, чтобы вам было удобнее работать в разных цветовых пространствах.
Asian Loto с виниловым покрытием предохранительной накладкой для блокировки, ALC-CHSV по 120 рупий за упаковку | Замки блокировки
О компании
Год основания 2015
Юридический статус Фирмы Частное лицо — Собственник
Характер бизнеса Производитель
Количество сотрудников От 11 до 25 человек
Участник IndiaMART с августа 2010 г.
GST06AEHPA0005 *
GST06AEHPA60005 Экспорт в Бангладеш
ASIAN LOTO Corporation является сертифицированной компанией ISO 9001: 2008 компанией, производителем, поставщиком и экспортером LOCKOUT TAGOUT и продуктов безопасности, которые имеют сертификат CE для обеспечения групповой изоляции и безопасного запирания и маркировки в соответствии с мировым стандартом безопасности OSHA.
Нашим мотивом является обеспечение определенной системы безопасных рабочих процедур реализации LOTO для сотрудников, подрядчиков и их персонала при работе по установке, техническому обслуживанию, очистке, производству, обработке или ремонту заводов или оборудования.
Большинство промышленных аварий вызвано либо техническими, либо человеческими причинами, которые являются незапланированной деятельностью, следовательно, это непредсказуемая и случайная причина, которая может произойти по множеству причин, таких как высвобождение накопленной энергии, небезопасная механическая конструкция и конструкция, неисправное устройство, неправильное обращение с материалами, протечки или неправильное использование инструментов и т. д.
Принимая во внимание, что этих несчастных случаев можно избежать и предотвратить с помощью надлежащих процедур LOCKOUT TAGOUT, небольшая неосторожность может привести к очень опасным несчастным случаям.
Мы предоставляем Полный ассортимент продукции LOCKOUT TAGOUT, который устанавливает ответственность работодателя по защите сотрудников от опасных источников энергии на машинах и оборудовании во время обслуживания и ремонта.
Стандарт OSHA’S LOCKOUT TAGOUT разработан для предотвращения вредного воздействия опасной энергии, потому что травмы в результате неспособности контролировать опасную энергию во время технического обслуживания могут быть серьезными или смертельными. , или перелом частей тела, и другие из-за несанкционированного или случайного использования энергии.
Требуется предотвратить появление опасного источника энергии, который включает электрические, сжатые газовые или гидравлические системы, заряженные пружины, нагретые вещества, радиацию, канализационные системы и накопленную гравитационную или кинетическую энергию.
Также обеспечение надлежащего обучения для руководства основным аспектом обучения заключается в руководстве программой управления энергопотреблением, элементами процедуры контроля энергопотребления, относящимися к обязанностям сотрудника, и различными требованиями, связанными с блокировкой / маркировкой.
Наша миссия заключается в изготовлении и поставке КАЧЕСТВЕННОЙ БЛОКИРОВКИ БЕЗОПАСНОСТИ И ПРОДУКТОВ безопасности, в том числе комплектов этикеток для предохранительной блокировки, навесных замков, аксессуаров для навесных замков, предохранительной блокировки, блокировки выключателя, блокировки предохранителя, блокировки задвижки, блокировки шарового клапана , Блокировка клапана-бабочки, Универсальная блокировка клапана, Аварийный выключатель / Блокировка кнопки, Многоцелевые коробки блокировки электрических панелей, Блокировка кабеля, Блокировка цилиндра, Блокировка шнура питания Plug, Пневматическая блокировка, Этикетки и бирки блокировки, Держатель разрешения, Безопасность блокировки, знаки и плакат , Учебное пособие по блокировке, Блокировка автомобиля, Дисплей стендов блокировки, Станция блокировки / шкаф (для ключей, навесных замков и LOTO), Сумка / мешочки с этикеткой для блокировки, Коробка для предложений, Блок групповой блокировки, Защитные ленты, Парковка и продукты для безопасности дорожного движения, Акустические уплотнения , Защита головы, Защита глаз, Защита ушей, Защита рук, Защита ног, Защита тела и поставщик других товаров для промышленной безопасности.
Кроме того, используя систему маркировки опасности и предостережения для идентификации, выполняемой с помощью замка для персонала, сотрудник должен рассмотреть возможность использования бирки опасности и удаления бирки опасности.
Когда каждый человек закончил работу, остановил работу в конце рабочей смены или уволился с работы, Знак опасности должен быть удален. Если использование оборудования небезопасно, необходимо прикрепить бирку о неработающем оборудовании, а также предоставить надлежащую информацию руководителю участка.